















Homeoboxové geny
Příroda je tu stále. Udržuje sama sebe.
José Ortega y Gasset, Vzpoura davů
V místě, kde cesta dělící Žlutý kopec od úbočí Špilberku opouští svahy obou brněnských vrchů, stojí starý klášter augustiniánů. Je jen málo tak významných míst, kde se utvářely základy moderní biologie. Pouze skromná výstavka připomíná, že zdejší opat Johann Gregor Mendel založil svou neskonalou pílí a přesným pozorováním nový vědní obor, genetiku. Třicet let po jeho téměř zapomenutých pokusech razil W. Bateson termín homeosis (ř. homeo - podobný). Označil jím biologické variace, při nichž se jeden článek segmentálně se opakujícího uspořádání struktur organizmu mění tak, že se podobá jinému. Později uveřejnil příklady homeózy z říše rostlin i živočichů a vysvětlil je genetickými variacemi. Ve dvacátých letech našeho století byly v Morganově laboratoři popsány homeotické geny, řídící tyto variace, na malé octomilce Drosophila melanogaster. Některé z nich dokonce byly identifikovány ve formě volných shluků na třetím chromozomu této mušky a byly zjištěny projevy jejich mutací.
Po druhé světové válce začal homeotické mutace sledovat (a činí tak dodnes) E. B. Lewis z Kalifornské techniky. Jedna z nich představovala monstrózní změnu zvanou Bithorax, kdy se vyvíjely mušky s jedním hrudním článkem nesoucím pár křídel navíc. Další byla mutace zvaná Antennapedia, která označovala jedince nesoucí na místě tykadel pár nožek. Lewis zjistil, že pro vznik monstra s dvěma páry křídel nebo s nohama na místě tykadel stačí mutace jednotlivých genů, i když samo vytvoření těchto abnormalit vyžadovalo aktivitu stovek dalších genů. Bylo jasné, že původní mutace musela proběhnout v nadřazených genech.
První důkazy
První homeotické geny, nazvané Ultrabithorax, Abdominal A a Abdominal B, izolovali D. S. Hogness a W. Bender na Stanfordově univerzitě začátkem osmdesátých let. Roku 1983 určili W. J. Gehring z Basilejské univerzity a W. McGinnis z Yaleovy univerzity v New Haven strukturu genu Antennapedia a objevili další homeotické geny tohoto komplexu. Zjistili také, že podobné geny se vyskytují u stonožek a dešťovek. Konzervativní struktura DNA byla nazvána homeobox. Tento exon o velikosti 183 nukleotidů kóduje tzv. homeodoménu, protein složený ze 61 aminokyselin. Funkcí této homeodomény je vazba na DNA podřízených genů, které jsou přímo ovládány homeoboxovými geny.Žáby a mouchy
Ve stejném roce se Gehringův spolupracovník E. J. De Robertis začal zajímat o to, zda se podobné geny vyskytují také u obratlovců. Jako model vybral africkou drápatku Xenopus laevis. Pohnutky, které ho k uskutečnění záměru vedly, popsal později takto: "Když jsme sledovali, jak ohromný pokrok byl vykonán studiemi na drozofile, bylo jasné, že kdybychom nalezli podobné řídící geny u obratlovců, získali bychom konečně srovnatelné vysvětlení zákonitostí jejich embryonálního vývoje [...] Byl pramalý důvod předpokládat, že žabí DNA bude takový gen obsahovat, nebo že by si geny tak příbuzensky vzdáleného druhu mohly být vzájemně podobné [...] Avšak stále jsme cítili, že to za to stojí [...] A brzy jsme oslavovali s lahví šampaňského [...]" (Scientific American, 263, 46, červenec/1990). Jaké bylo pak překvapení, když se dále zjistilo, že stejné geny odpovídají za výstavbu těla drozofily i žáby a že je možné protilátkami proti homeodoménové bílkovině změnit u pulce vývoj přední míchy v zadní mozek.
Pohnutky, které ho k uskutečnění záměru vedly, popsal později takto: "Když jsme sledovali, jak ohromný pokrok byl vykonán studiemi na drozofile, bylo jasné, že kdybychom nalezli podobné řídící geny u obratlovců, získali bychom konečně srovnatelné vysvětlení zákonitostí jejich embryonálního vývoje [...] Byl pramalý důvod předpokládat, že žabí DNA bude takový gen obsahovat, nebo že by si geny tak příbuzensky vzdáleného druhu mohly být vzájemně podobné [...] Avšak stále jsme cítili, že to za to stojí [...] A brzy jsme oslavovali s lahví šampaňského [...]" (Scientific American, 263, 46, červenec/1990). Jaké bylo pak překvapení, když se dále zjistilo, že stejné geny odpovídají za výstavbu těla drozofily i žáby a že je možné protilátkami proti homeodoménové bílkovině změnit u pulce vývoj přední míchy v zadní mozek.
Architekti a stavitelé
Homeoboxové geny jsou po aktivaci přepsány na mRNA, která pak určuje homeodoménovou bílkovinu. V časných embryích jsou postupně aktivovány podél předozadní osy. Jejich funkce spočívá v precizní časoprostorové regulaci vývoje plodu, respektive v jejich postupném zapojování v přesném pořadí.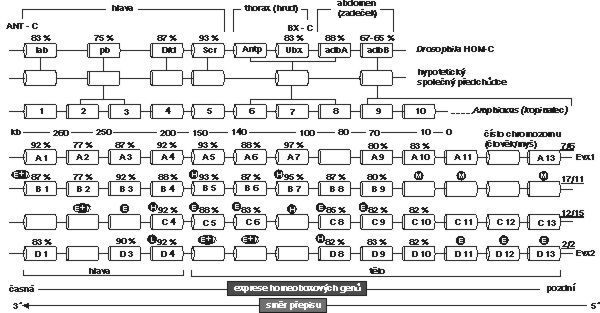
Je již naprosto jasné, že homeoboxové geny určují základní tělesný plán nejen bezobratlých živočichů, ale i obratlovců. Na základě dnes dostupných informací o homeoboxových genech si můžeme představit embryo rozdělené do jednotlivých buněčných polí orientovaných v průběhu podélné osy a odlišujících se rozdílnou schopností vývoje (viz obrázek). Přesně určené okrsky se stanou určitými tkáněmi a orgány. Pokusy na transgenních myších (ovlivněných umělým vložením genu) ukázaly, že zničení příslušných homeoboxových genů vedlo k vývojovým změnám stavby těla (pozn. red.: podobný význam mají homeogeny při určování prstů a částí končetin, viz např. Vesmír 74, 76, 1995/2).
Nejvíc udivuje již zmíněný vysoký stupeň konzervativnosti homeoboxových genů. Lidský HOXA7 se liší pouze v jedné ze šedesáti pozic od homeodomény Antp (viz obrázek), tj. od doby oddělení předků členovců a strunatců před více než 600 miliony let. Všichni čelistnatí obratlovci mají čtyři homeoboxové komplexy, z nichž každý je lokalizován na jiném chromozomu. Je pravděpodobné, že tyto komplexy vznikly v průběhu evoluce duplikací jediné sady homeoboxových genů, což je stav typický pro bezobratlé živočichy. Nasvědčují tomu i nedávno provedené studie u kopinatce. Tento mořský živočich je archaickým zástupcem původních strunatců a unikátní připomínkou klíčového mezistupně vývoje homeotických genů. Jejich uspořádání v genomu kopinatce odpovídá hypotetickým předpokladům, jak asi musel vypadat stav u předků obratlovců i segmentovaných bezobratlých živočichů. Genom kopinatce má totiž jen jedinou sadu homeoboxových genů, které jsou homologické nejméně prvním deseti paralogovým skupinám Hox-genů obratlovců, dokonce ve stejném pořadí.
Kontroloři růstu
Od konce osmdesátých let rychle vzrůstá počet prací zabývajících se projevem homeoboxových genů a funkcí homeodoménových bílkovin regulujících buněčnou diferenciaci. Intenzivní studium těchto otázek zasahuje přímo do jedné z nejatraktivnějších oblastí současné imunologie, do diferenciace imunitních buněk a krvetvorby, včetně vzniku jejich poruch. Např. z homeoboxových genů třídy I byla popsána účast HOXA1, HOXA10, HOXB6 a HOXC6 u lidských nádorových onemocnění. U myší vede neadekvátní exprese genu Hoxb8 v buňkách kostní dřeně k leukemii. U beztymových myší způsobuje tento gen maligní zvrhnutí fibroblastů ve fibroblastom. K malignímu zvratu vede i nepřítomnost genu Hoxd3.Sirotčí geny
Kromě homeotických genů uspořádaných do jednotlivých sad (genů třídy I) byly identifikovány takzvané sirotčí geny, které jsou v savčím genomu umístěny mimo tyto sady. Jsou to například oct-geny octamerové rodiny, evx-rodiny (z homologie even-skipped homeoboxových genů drozofily) a en-rodiny (homologní engrailed genům drozofily) a řada dalších individuálních genů.
Nemluvě o řadě sirotčích genů zasahujících do morfogenezy mozku, metanefru nebo končetin byla prokázána jejich důležitost pro plnohodnotné fungování imunitního systému. Lze uvést např. geny Oct-2 a Pit I, jejichž přítomnost je dokonce pokládána za nezbytnou při tvorbě protilátek plazmatickými buňkami. Z tohoto pohledu rozvířil nedávno zájem vědecké veřejnosti jiný sirotčí gen, gen HOX11, který byl původně izolován při akutní lymfoblastické leukemii. Tento gen se nachází v blízkosti genu pro antigenní receptor T-lymfocytů a jeho homeodoména je totožná s produktem myšího genu Hox11. Roku 1994 prokázal Robertsův tým z Lékařské fakulty Univerzity v St. Louis jeho význam při genezi důležitého orgánu imunity - sleziny. Studie s tzv. knokautovanými myšmi (míněny myši s uměle zničeným genem Hox11), jež neměly slezinu, poprvé dokázaly, že homeoboxové geny nesouvisí jen s osovým uspořádáním organizmu, ale jsou rovněž odpovědné za vznik orgánů, a navíc jediný homeoboxový gen může určovat vznik a vývoj jediného orgánu.
Geny z prahor
Homeotické geny byly nalezeny téměř u všech zástupců mnohobuněčných živočichů i rostlin a každý týden přináší nové objevy. Bylo by zajímavé uvažovat o prvních mechanizmech umožňujících přechod od kolonií, které se skládají z ještě poměrně samostatných jednobuněčných organizmů, ke složitě řízeným státům eumetazoí, mnohobuněčných jedinců. Víme, že svou roli sehrály bílkoviny mezibuněčné hmoty jako kolageny, fibronektin, vitronektin, proteoglykany, SPARC, tenascin nebo laminin a řada dalších, které jsou evoluční novinkou starou jen asi 700 milionů let. Genetickou řídící informaci pro strukturální a fyzické udržení integrity však nesly homeotické geny. Proto se uplatňují i v řídících soustavách organizmu - nervové, hormonální a imunitní. Ze stovek homeotických bílkovin pouze nepatrná část řídí výstavbu živočicha podle předozadní osy, a právě tato velmi konzervativní skupina byla objevena jako první. Mnohočetnému výskytu homeotických genů nasvědčují nedávné poznatky Japonců o živočišných sladkovodních houbách. Japonští vědci dokonce předpokládají, že tito živočichové s jednoduchou stavbou těla mohou mít více homeoboxových genů, než mají buněčných typů. To by znamenalo, že tyto geny nejsou angažovány jen v procesech provázejících strukturální organizaci.Při přechodu od základního tělesného plánu strunatců k výstavbě těla obratlovců se celý shluk homeoboxových genů několikrát zdvojil a jistě se geny i specializovaly (mimochodem je dnes známo už více než 100 homeoboxových genů u savců a více než 50 u drozofily). To mělo za následek posun řídících funkcí v rámci základního tělesného plánu. Byla vytyčena cesta ke vzniku nových struktur a orgánů, směr, který už nadále vedl ke zvyšování složitosti organizmů. Tomuto předpokladu nasvědčují pokusy s odstraněním genu Hoxa2 u myší. Je obecně známým faktem, že savčí sluchový orgán se vyvinul z primitivní čelistní kosti, kterou mají ještě plazi. V nepřítomnosti genu Hoxa2 se zárodky myší vyvíjejí s pozměněnými kostními elementy, které připomínají stav u plazů.
Pro futuro
Objev homeotických regulací (zatím víceméně nedoceněný) ovlivní biologii 21. století. Jeho následky zatím nemůžeme ani tušit. Nelze pominout ani fakt, že výskyt regulačních homeotických genů téměř u všech eukaryontních organizmů znamená definitivní potvrzení jednoty života na Zemi. Objevem homeotických genů se otevřela kapitola té nejvíce vzrušující oblasti genetiky, jejímž cílem je nejen porozumět, ale také se naučit ovlivňovat procesy, v jejichž důsledku se z oplozeného vajíčka zrodí složitý organizmus. Pochopení, proč je zpočátku tolik podobný vývoj různých organizmů zakončen zrodem neopakovatelného jedince, jistě přispěje k dalšímu objasnění hry přírody, kterou nazýváme evoluce. Na této cestě poznání se "homeobox může stát biologickým ekvivalentem rosetské desky", jak výstižně shrnuli význam těchto genů G. Struhl a J. Slack už v době jejich objevu (G. Struhl, Nature
Jak se dělá oko?
 Pod názvem "O evoluci očí: je libo jednoduché, nebo složené?" komentuje C. S. Zuker v loňském srpnovém čísle časopisu Science izolaci genu eyeless, odpovědného za vývoj složeného oka u drozofily. Tento gen je homologem genu Pax-6 z malé rodiny genů, které jsou aktivní zejména v embryonálních základech nervové trubice a základech oka. Tyto geny byly nalezeny u všech tříd obratlovců a jsou ještě konzervativnější než homeoboxové geny Hox. Nově izolovaný gen Pax-6 vykazuje z 94 % identitu mezi drozofilou a člověkem. U heterozygotních jedinců, kteří mají pouze jednu alelu genu ze dvou, chybí duhovka (syndrom Aniridia u lidí) a zmenšuje se velikost očí (Small eye u hlodavců). Příbuzné struktury DNA jako u genu Pax-6 byly zjištěny polymerázovou řetězovou metodou u ploštěnek, pásnic a měkkýšů.
Pod názvem "O evoluci očí: je libo jednoduché, nebo složené?" komentuje C. S. Zuker v loňském srpnovém čísle časopisu Science izolaci genu eyeless, odpovědného za vývoj složeného oka u drozofily. Tento gen je homologem genu Pax-6 z malé rodiny genů, které jsou aktivní zejména v embryonálních základech nervové trubice a základech oka. Tyto geny byly nalezeny u všech tříd obratlovců a jsou ještě konzervativnější než homeoboxové geny Hox. Nově izolovaný gen Pax-6 vykazuje z 94 % identitu mezi drozofilou a člověkem. U heterozygotních jedinců, kteří mají pouze jednu alelu genu ze dvou, chybí duhovka (syndrom Aniridia u lidí) a zmenšuje se velikost očí (Small eye u hlodavců). Příbuzné struktury DNA jako u genu Pax-6 byly zjištěny polymerázovou řetězovou metodou u ploštěnek, pásnic a měkkýšů.Po objevu homeoboxových genů není tak překvapující, že vývoj oka drozofily a člověka je řízen homologním genem, protože zástupci obou živočišných skupin sdílejí ve vývoji řadu společných molekulárních mechanizmů.
Je-li např. lidský HOXD4 (dříve Hox4B) přenesen do embrya drozofily, funguje jako gen Dfd. Je však zarážející, že diferenciace orgánů tak anatomicky rozdílných u much a lidí je kontrolována celou kaskádou subordinovaných homologických genů.
Optické orgány schopné zobrazovat se nacházejí nejméně u šesti živočišných kmenů (žahavci, měkkýši, kroužkovci, členovci, drápkovci a strunatci). Avšak všechny, i ty nejjednodušší (například u ploštěnek) využívají k záchytu fotonů stejné receptorové molekuly a univerzální systém přenosu signálu, který mohl být řízen v dávných dobách právě předchůdcem genu Pax-6. Bude-li podobný princip homologické genetické kontroly potvrzen také u jiných struktur pokládaných za analogické, budeme muset naše dosavadní názory na tradiční pojetí vztahu analogie versus homologie v evoluci živočišných taxonů podrobit důkladné revizi.
Pigmentové světločivné skvrny jsou známy u celé řady druhů prvoků, jako je např rod Euglena, krásnoočko. Nejrůznější typy očí se pak vyskytují u nejrůznějších skupin živočichů, počínaje těmi téměř nejprimitivnějšími. Málo známé jsou oči žahavců (Cnidaria), kterými jsou "vybaveny" např. medůzky rodu Charybdaea. Naopak všeobecně jsou známé "primitivní" oči ploštěnek, jejichž počet a rozmístění na hlavovém konci těla se v mnoha případech "používá" i jako určovacího znaku. Také o očích kroužkovců se toho mnoho neví. Je však pozoruhodné, že zde nalézáme jak oči složené, tak jednoduché s jedinou čočkou. Stejně je tomu i u členovců - zatímco kupříkladu pavouci (Tegenaria domestica - pokoutník domácí) mají jednoduché oko s jedinou čočkou, rohovkou a sítnicí, oči hmyzu jsou složené z mnoha drobných oček z nichž každé má vlastní krystalinní tělísko (čočku) i vlastní pigmentové a senzorické buňky. Pozoruhodná je stavba očí u některých měkkýšů, a to u plžů (např. Helix pomatia - hlemýžď zahradní) i u mlžů (např. rodu Pecten - hřebenatka). Hřebenatka má ještě "buněčnou čočku", což je považováno za původní, primitivní či jednodušší stav. Nejdokonalejší stavbu však mají tzv. komorové oči hlavonožců (Octopus - chobotnice).
O autorech
Petr Šíma
Ilja Trebichavský
Doporučujeme

