









Tolerance pylu vůči vyšším teplotám
| 4. 11. 2024V důsledku klimatické změny čelí zemědělství vážným výzvám. Zvyšující se teploty mají přímý dopad na morfologii a fyziologii rostlin, což způsobuje pokles výnosů a ohrožuje potravinovou bezpečnost. Teplotní stres zasahuje pylová zrna, která jsou nejcitlivější částí rostliny. Výsledkem jsou poruchy opylení a plodnosti, které mohou vést k zásadním ekonomickým ztrátám v zemědělství.
Rostliny zažívají teplotní stres po překročení prahové teploty, většinou 5–10 °C nad optimum, přičemž jejich reakce závisí na délce a intenzitě stresu. I mírnější výkyvy však mohou mít významné ekonomické dopady. Například zvýšení minimální teploty o 1 °C v období růstu způsobuje pokles výnosů rýže (Oryza sativa) o 10 %. Podobně zvýšení denních i nočních teplot o pouhý 1 °C nad optimum vede ke snížení výnosů pšenice (Triticum aestivum) o 5 %.
Zvýšené teploty citelně zasahují i proces rozmnožování rostlin, což vede k snížení plodnosti mnoha druhů. Pohlavní rozmnožování krytosemenných rostlin zahrnuje tři fáze: vývoj reprodukčních orgánů, opylení a růstu pylové láčky a konečně vývoj embrya a semene. Teplotní stres zasahuje samčí a samičí pohlavní orgány rozdílně, přičemž pyl je během svého vývoje vůči působení vysokých teplot citlivější. Teplotní stres ovlivňuje množství a morfologii pylu, strukturu buněčných stěn a jeho celkový metabolismus.
Vliv teplotního stresu na pyl
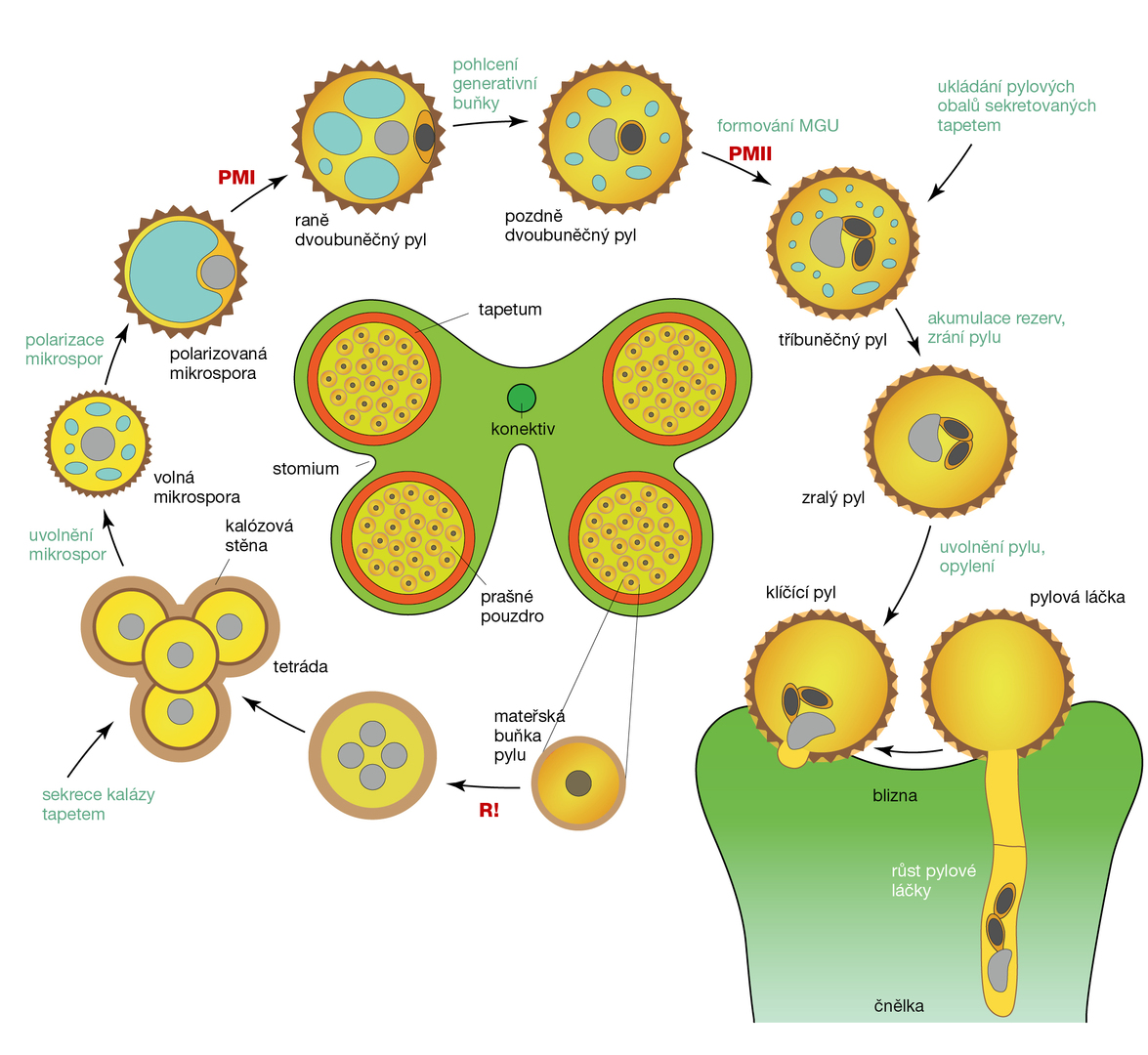
Diploidní pylové mateřské buňky (mikrosporocyty) podstupují uvnitř prašníků meiotické dělení, čímž vznikají čtyři haploidní mikrospory. Ty drží pohromadě v tetrádě silnou stěnou tvořenou polysacharidem kalózou. Kalózová stěna je později rozložena kalázou, enzymem vylučovaným buňkami tapeta. (Tapetum je výstelková vrstva metabolicky velice aktivních buněk odpovědných za výživu vyvíjejících se pylových zrn.) Tím se uvolní jednotlivé mikrospory. Od tohoto okamžiku mikrospory samostatně rostou, vakuolizují se a jejich jádra putují k buněčnému okraji. Polarizované mikrospory následně procházejí vysoce asymetrickým mitotickým dělením zvaným první pylová mitóza (PMI), při kterém vzniká velká vegetativní buňka a malá generativní buňka. Generativní buňka podstupuje ještě jedno mitotické dělení, druhou pylovou mitózu (PMII), za vzniku dvou spermatických buněk, samčích gamet (obr. 1).
Úspěšné a koordinované opylení a oplození vyžaduje synchronizovaný vývoj mikrospor uvnitř prašníku. Tento proces je složitě řízen, a pokud selže (např. kvůli stresu), asynchronní vývoj vede k fyziologickým a metabolickým rozdílům mezi mikrosporami. U většiny rostlin se zdá, že právě nástup meiózy a vývoj mikrospor směrem k první pylové mitóze je nejvnímavějším obdobím vůči působení stresových podmínek.
Nejčasnější vývojové defekty pylu u zkoumaného huseníčku rolního (Arabidopsis thaliana) vyvolané teplotním stresem skutečně nastávají během meiózy, kdy se zvyšuje frekvence rekombinačních událostí. Teplotní stres ovlivňuje také dynamiku cytoskeletu a orientaci dělicího vřeténka. Způsobuje nárůst množství některých metabolitů, konkrétně ochranných molekul. Kupříkladu ve stresovaných polarizovaných mikrosporách rajčat (Solanum lycopersicum) stoupá koncentrace flavonoidů a polyaminů, které hrají důležitou roli při detoxikaci reaktivních forem kyslíku (ROS).
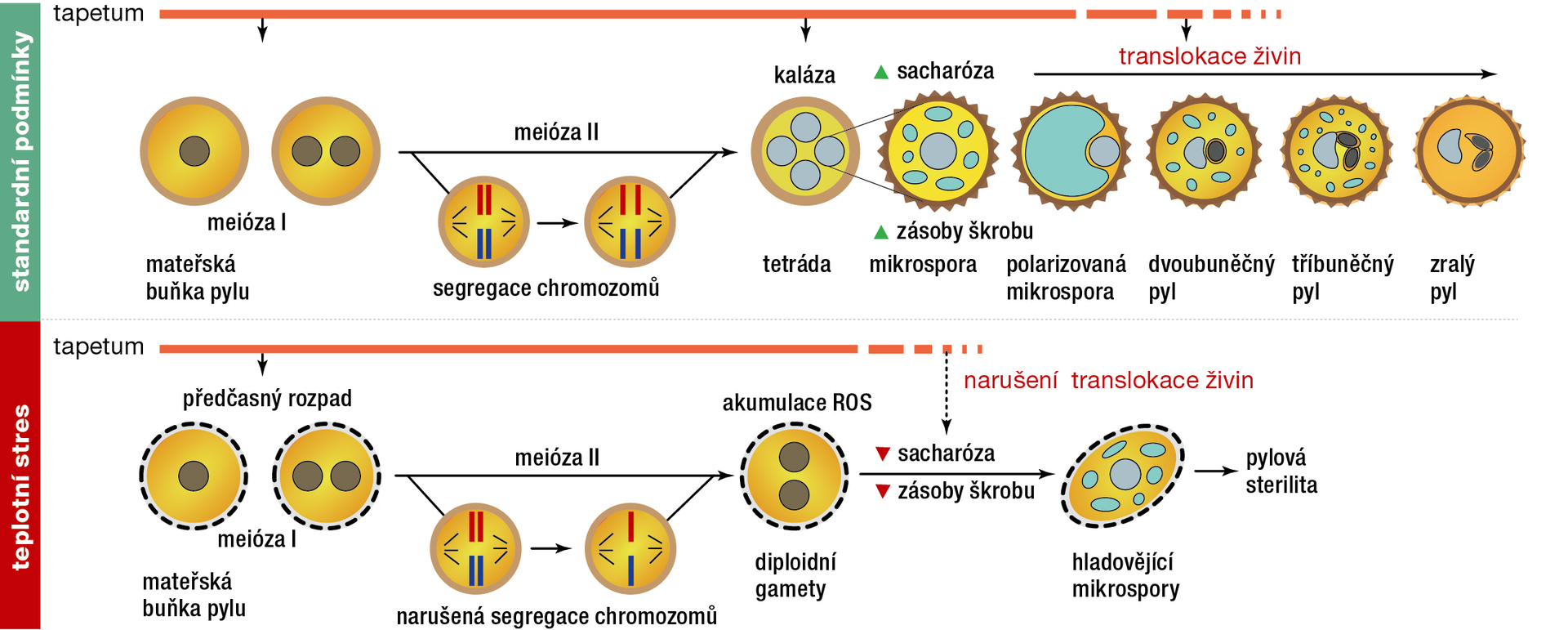
Teplotní stres způsobuje předčasnou degradaci tapeta. Tapetum je značně bohaté na mitochondrie. Právě ty přispívají během působení teplotního stresu k dramatickému nárůstu ROS, vedlejších produktů aerobního metabolismu mitochondrií. Hromadění ROS způsobuje oxidační poškození a může vést k buněčné smrti. Během teplotního stresu skutečně dochází k hromadění ROS v prašnících, což způsobuje nerovnováhu mezi hladinami ROS a enzymy, které je neutralizují. To pak vyvolává předčasnou degradaci vrstvy tapetálních buněk, která je příčinou samčí sterility (obr. 2). Reaktivní formy kyslíku však také spouštějí ochranné mechanismy rostlinných buněk. Kupříkladu indukují expresi klíčového proteinu HEAT SHOCK TRANSCRIPTION FACTOR A1 (HsfA1). Tento základní člen rodiny transkripčních faktorů aktivovaných stresem (HSF) dále stimuluje expresi genů odpovědných za reakci na teplotní stres.
Při opylení pylové zrno dopadne na bliznu, kde rehydratuje a vyklíčí v pylovou láčku. Pylové láčky jsou specializované struktury, které dopravují samčí pohlavní buňky do zárodečného vaku pro dvojité oplození. Jsou také nejrychleji rostoucími rostlinnými buňkami (Vesmír 99, 685, 2020/12). Jejich růst je proto energeticky náročný proces, který závisí na ukládání a využívání zásob, rychlé syntéze buněčné stěny a přesné komunikaci mezi buňkami. Degradace mitochondrií při vysokých teplotách způsobuje poruchy těchto procesů, a tím i zpomalení jejich růstu, jak bylo pozorováno u pylu rýže a pylových láček rajčat.
Reakce na teplotní stres
Rostlinné buňky mohou rozpoznat působení stresu na několika úrovních pomocí specifických senzorů. Ty zahrnují fosfolipidové membrány (jejich fluiditu a propustnost), hladiny vápenatých iontů, stabilitu proteinů, detekci nesprávně složených proteinů a odpověď na ně (Unfolded Protein Response, UPR), stav chromatinu a modifikace histonů či strukturu a stabilitu mRNA. Jednotlivé stresové senzory spouštějí signální kaskády, které vyvolávají reakce na teplotní stres a umožňují obnovení buněčné homeostázy.
Hromadění nesprávně složených nebo nesložených proteinů v endoplazmatickém retikulu aktivuje evolučně značně konzervovanou UPR dráhu. U rostlin byly popsány dvě její větve a obou se účastní transkripční faktory velké genové rodiny bZIP. První zahrnuje nekonvenční sestřih transkriptů kódujících protein bZIP60, druhá zahrnuje dvojici transkripčních faktorů ukotvených v membráně, bZIP17 a bZIP28. Ty jsou při teplotním stresu uvolněny a přeneseny do jádra, kde aktivují genovou expresi. V pylu je za teplotního stresu aktivní pouze první větev UPR dráhy, ačkoli během vývoje pylu byla detekována exprese genů účastnících se obou větví UPR.
Hromadění nesprávně složených proteinů v cytoplazmě spouští UPR prostřednictvím komplexů proteinů teplotního šoku (HSP, z ang. heat shock proteins) a transkripčních faktorů teplotního šoku (HSF). Již zmíněný protein HsfA1 funguje jako hlavní regulátor aktivace odpovědi na teplotní stres ve vegetativních pletivech, je však aktivní i v reprodukčních orgánech, kde mimo jiné kontroluje expresi dalšího člena rodiny HSF, proteinu HsfA2. Právě ten reguluje v pylu rajčat skupinu proteinů teplotního šoku, která hraje při zmírňování účinků teplotního stresu na metabolismus rostlin klíčovou roli. Tyto proteiny působí převážně jako molekulární chaperony,1) které chrání ostatní proteiny. Potlačení exprese HsfA2 logicky snižuje životaschopnost pylu a jeho klíčivost, ale jen během časného vývoje pylu. V pozdějších fázích již neúčinkuje. To ukazuje, že HsfA2 je důležitým hráčem v procesu aklimatizace. Podporuje termotoleranci pylu a chrání jej i v pozdějších fázích vývoje.
Moderní přístupy
Reakce rostliny na teplotní stres se projevuje hierarchickým přeprogramováním genové exprese a buněčného metabolismu na všech úrovních – transkriptomu (souboru všech RNA přepsaných z DNA), translatomu (souboru mRNA, které jsou na ribozomech „překládány“ do proteinů), proteomu (souboru proteinů) i metabolomu (všech malých molekul produkovaných buněčným metabolismem. Díky pokroku v izolačních i detekčních technikách jsme nyní schopni studovat různé aspekty těchto reakcí v jednotlivých buňkách a buněčných typech účastnících se rozmnožování. Jedná se např. o pylová zrna, pylové láčky, spermatické buňky či vajíčka.
Profilování transkriptomu dokáže identifikovat aktivní geny přítomné ve studovaném vzorku v daném čase. K němu se rutinně používá sekvenování RNA (RNA-seq), které postupně nahradilo dříve používané genové čipy. RNA-seq nabízí širší dynamický rozsah, vyšší citlivost a pokrytí celého genomu, což vede k popsání neznámých transkriptů a nových sestřihových variant. Hlavní výhodou RNA-seq je však jeho aplikovatelnost na nemodelové druhy, umožněná neustálým vývojem výpočetních nástrojů pro integraci dat z různých studií využívajících různé platformy a metody.
U huseníčku bylo prokázáno, že teplotní stres mění přibližně 15 % pylového transkriptomu. Ranější vývojové fáze jsou na teplotní výkyvy opět citlivější než fáze pozdější. Transkriptomické studie také odhalily regulaci exprese genů na posttranskripční úrovni, konkrétně prostřednictvím alternativního sestřihu. Ten tedy pravděpodobně hraje důležitou roli v adaptaci na teplotní stres. Svědčí pro to i výzkum pylu rajčat, v němž mezi alternativně sestříhané transkripty patřily i ty kódující HSF a HSP.
Vztah mezi transkripční a translační regulací je možno zkoumat srovnáním transkriptomických a translatomických dat. Analýza translatomu ukazuje na celkově vysokou korelaci mezi transkripčními a translačními odpověďmi na zvýšené teploty, nicméně významná část genů vykazuje specifickou formu regulace na translační úrovni. Posttranskripční regulace a reorganizace ribozomů během stresové reakce zahrnuje řízenou redistribuci mRNA mezi aktivně překládajícími ribozomy a cytoplazmatickými granulemi mRNA, v nichž jsou skladovány zásobní transkripty, jejichž translace je blokována.
Několik studií využívajících imunoprecipitaci chromatinu prokázalo zapojení epigenetických procesů do stresové reakce v pylu. Teplotní stres například mění konformaci chromatinu a ovlivňuje metylaci DNA, konkrétně například genů kódujících HSP. Podobně byla popsána i důležitost malých nekódujících RNA, které se angažují v reakci na teplotní stres specificky podle vývojové fáze, a to na transkripční či epigenetické úrovni.
Buněčný proteom plně neodpovídá transkriptomu, a to zejména v systémech s vysokou mírou translační regulace, jakým je právě pyl. Proto je nutné doplňovat transkriptomiku i translatomickými a proteomickými daty. Nízká korelace mezi hladinami transkriptů a proteinů během reakce na teplotní stres byla pozorována například u pylu rajčete. Vzhledem k výše popsané podobnosti transkriptomu a translatomu ve stresovaném pylu huseníčku se zdá, že se na této regulaci podílejí i posttranslační úrovně. Je zajímavé, že teplotní stres u pylu rajčete ukázal nejen oddělené reakce na transkripční a posttranskripční úrovni, ale také neobvykle vysoký podíl pylově specifických transkriptů a proteinů mezi takto regulovanými transkripty.
Rostlinný metabolom je velmi komplexní, protože zahrnuje produkty jak primárního, tak sekundárního metabolismu. Odhaduje se, že rostliny obsahují až 200 000 různých metabolitů. Bohužel hlavním omezením v profilování metabolomu je absence jednotné metodiky, neboť jednotlivé metabolity se liší z hlediska koncentrace a chemických vlastností. Velkou analytickou výzvou, pokud jde o pyl, je také odstranění pylových obalů a dalších hydrofobních sloučenin na povrchu pylového zrna, které brání detekci a identifikaci různých metabolitů.
Teplotní stres vyvolává metabolickou nerovnováhu. V tomto ohledu je akumulace reaktivních forem kyslíku spolehlivým ukazatelem stresu a souvisí se značným poklesem jeho klíčivosti. Zajímavá je také koordinovaná úloha rostlinných hormonů a sacharidů ve stresové reakci. Několik studií ukázalo silnou interakci mezi drahami zprostředkovanými sacharidy a kyselinou abscisovou (ABA). ABA potlačuje v pylu pšenice expresi invertázy (enzymu štěpícího sacharózu), což vede k narušení metabolismu cukrů. Podobně byla popsána regulační interakce mezi kyselinou jasmonovou a metabolismem sacharidů, což má vliv na transport vody do prašníků. Aplikace auxinu vedla ke zvýšení termotolerance reprodukčních orgánů rýže a ke snížení samčí sterility.
Přes nepochybné úspěchy jsme zatím pouze na začátku poznávání metabolické regulace. Komplexní profilování metabolitů za stresových podmínek a zasazení získaných výsledků do souvislostí s výsledky transkriptomických a proteomických studií bude dalším nezbytným krokem.
Mechanismy termotolerance
Negativní účinky teplotního stresu lze do jisté míry obejít, pokud rostliny projdou aklimatizací. Základní termotolerance představuje schopnost rostliny odolat mírnému teplotnímu stresu. Následuje-li krátká fáze zotavení, indukuje získanou termotoleranci, která umožňuje rostlinám odolat většímu, jinak smrtícímu teplotnímu stresu. Tento jev lze přičíst schopnosti rostlinných buněk ukládat proteiny, které mohou zvýšit jejich termotoleranci. Například rajčata vykázala dramatický pokles klíčivosti pylu, když byla dvě hodiny vystavena 50 °C. Aklimatizace následovaná fází zotavení však vedla ke zvýšené toleranci ve srovnání s rostlinami, které takto ošetřeny nebyly. Taková aklimatizace není specifická jen pro pyl, ale podobně ovlivňuje i vývoj semen. Další studie na rajčatech ukázaly, že pylovou termotoleranci lze zvýšit působením hormonů. Předběžné ošetření rajčat etylenem významně zvýšilo kvalitu pylu po působení stresu. Pylový proteom poté ukázal hojné zastoupení proteinů, které se podílejí na udržování buněčného redoxního stavu, což pravděpodobně minimalizovalo účinek vysokých teplot.
Termotolerance pylu je stále důležitější a má značný ekonomický význam, při šlechtění nových odrůd je proto třeba jí věnovat pozornost. Je nezbytné vyvinout robustní protokoly, které umožní hodnocení kvality pylu v měnících se environmentálních podmínkách, a vytvořit odpovídající metodiku předběžného ošetření a aklimatizace plodin rostoucích v ohrožených oblastech. Výkyvy teplot zřídka představují ojedinělou událost. Teplotní stres často souvisí s vysycháním půdy a vyšší intenzitou osvětlení. Odpověď pylu na několik souběžně probíhajících stresů se může výrazně lišit od jeho reakce na samotný teplotní stres. To bylo prokázáno například u tabáku. Ve snaze simulovat realistické podmínky byly rostliny vystaveny buď jednotlivým stresům (teplu nebo suchu), nebo oběma současně. Na kombinovaný stres reagovaly odlišně, byly aktivovány jiné skupiny genů. Proto je třeba navrhovat experimenty napodobující skutečné podmínky. Moderní genomické nástroje mohou být v tomto ohledu velmi užitečné.
Většina dosavadních výzkumů věnovaných tepelnému stresu pylu však byla založena na transkriptomice a proteomice. Studium metabolomiky a dalších specializovanějších omických přístupů prozatím zaostává. Výzkumná data získaná během různých experimentů je proto třeba propojit tak, aby poskytla šlechtitelům cílené informace o genech, které mohou využít ke zlepšení genetických zdrojů a šlechtění odolných linií.
Poznámky
1) Chaperony jsou proteiny, které se starají o správnou prostorovou strukturu ostatních proteinů.
Ke stažení
 článek ve formátu pdf [657,64 kB]
článek ve formátu pdf [657,64 kB]
O autorovi
David Honys

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
