










DNA stále trochu záhadná
| 3. 4. 2023Pětadvacátý duben 1953 připadl na sobotu a nové číslo časopisu Nature vyšlo v Londýně jako obyčejně. Mezi jinými články našli čtenáři také tři krátké příspěvky pod souhrnným titulkem „Molekulová struktura nukleových kyselin“. Mezi nimi i ten nejslavnější, od Jamese Watsona a Francise Cricka [1]. Ani po sedmdesáti letech však struktuře dvoušroubovice v úplnosti nerozumíme.
„Vše, co dělají živé organismy, je možné pochopit na základě pohybujících se a kmitajících atomů,“ psal Richard Feynman ve svých Přednáškách z fyziky, které poprvé vyšly roku 1963. Epochálnímu objevu struktury DNA bylo tehdy právě deset let a biologie se probouzela do nové éry – období stále intenzivnějšího úsilí o pochopení podstaty života na atomární úrovni.

Navržená struktura DNA byla však zpočátku přijímána dosti zdrženlivě. Počet článků v časopise Nature, které pojednávaly o DNA, mezi léty 1950 a 1960 sotva překročil dvacet ročně a po uveřejnění Watsonovy a Crickovy práce se nijak nezvýšil. Články, které přímo zmiňovaly dvoušroubovici, se počítaly na jednotky. A to přesto, že role DNA jako dědičného materiálu byla v té době díky novým pokusům už mnohem jasnější. „Neušlo naší pozornosti,“ psali Watson a Crick, „že specifické párování, které zde postulujeme, okamžitě ukazuje na možný mechanismus kopírování genetického materiálu.“ Samotní autoři ale jen o měsíc později v jiném článku přiznávají, že dosud nic nenaznačuje, jak by takový mechanismus mohl fungovat.
„DNA není zdaleka tak statická a stejnorodá, jak by se mohlo zdát. Jde o dynamickou, polymorfní strukturu, jejíž konformační vlastnosti jsou ovlivněny sekvencí bází, které ji tvoří.“
Ani princip syntézy proteinů tehdy ještě nebyl znám. Teprve v roce 1957 formuloval Crick proslulé „centrální dogma“, podle nějž biologická informace směřuje od nukleových kyselin k proteinům, a první kodon – trojici bází kódujících aminokyselinu – určili Marshall Nirenberg a J. Heinrich Matthaei až roku 1961. Ani Feynman při psaní své knihy ještě princip kódování aminokyselin bázemi v DNA neznal.
Byl tu však i další problém – struktura dvoušroubovice v té podobě, v jaké ji Watson a Crick navrhli, jednoduše postrádala solidní experimentální základ. Byla zpočátku spíše geniální hypotézou než vědeckým faktem. Ve svých klíčových rysech byla potvrzena, mnohé však také bylo korigováno nebo zcela opuštěno. Připomeňme si ve stručnosti historii objevu i některé současné problémy týkající se struktury DNA.

Jak to všechno začalo
V úvodu zmíněný článek Jamese Watsona a Francise Cricka z Cavendishovy laboratoře v Cambridgi navrhoval strukturu DNA ve tvaru pravotočivé dvoušroubovice, s vlákny cukrfosfátové páteře obíhajícími po obvodu v opačných směrech a s bázemi uvnitř. Báze obou řetězců se v ní spojují vodíkovými můstky do párů, a to vždy pouze adenin (A) s thyminem (T) dvěma můstky a guanin (G) s cytosinem (C) rovněž dvěma. Jednotlivý řetězec může nést jakoukoli sekvenci bází. Je-li však již dána, určuje toto párování jednoznačně sekvenci druhého řetězce. Roviny párů jsou kolmé k ose dvoušroubovice a na jednu otočku jich připadá deset. Crickova žena Odile doprovodila článek vkusnou ilustrací (obr. 1A).
Další dva příspěvky v Nature z 25. dubna 1953 nabízely experimentální výsledky, těsně svázané se strukturou dvoušroubovice – autorem prvního byl Maurice Wilkins se spolupracovníky, druhý napsala Rosalind Franklinová se svým studentem [2]. Všichni pracovali na King’s College v Londýně.
Autoři těchto tří článků se navzájem znali a měli povědomí i o svých výsledcích. Zejména mimořádně kvalitní rentgenová fotografie (obr. 5) z laboratoře R. Franklinové, kterou Wilkins bezelstně ukázal Watsonovi při jeho návštěvě King’s v lednu 1953, nesla jednoznačné rysy helikální struktury. Watson s Crickem později přiznávali, že bez znalosti výsledků Rosalind Franklinové, předčasně zesnulé roku 1958, by nedokázali strukturu DNA v prvních měsících roku 1953 objevit.
Pan Hoogsteen a ti druzí
V následujícím období se vědci snažili prověřit různé aspekty této prvotní struktury. Zatímco obecné rysy již nevzbuzovaly pochybnost, detaily Watsonova-Crickova modelu, včetně specifického uspořádání vodíkových můstků v párech bází, se stále nedařilo potvrdit. Báze se totiž mohou párovat různými způsoby (do můstků se mohou zapojit různé atomy) a způsob navržený Watsonem a Crickem rozhodně není jediný možný.
Jeden směr výzkumu spočíval v získání krystalů izolovaných párů s nadějí, že zjištěné uspořádání bude odpovídat situaci v samotné dvoušroubovici. První takovou studii párů A–T publikoval Karst Hoogsteen roku 1959. Namísto Watsonova-Crickova párování však pozoroval jiný typ párů, dnes nazývaných jeho jménem, v nichž je purinová báze, tedy A nebo G, otočena o 180 stupňů kolem vazby, jež ji spojuje s cukrfosfátovou páteří (obr. 2). V roce 1963, rok poté, co Watson a Crick spolu s Wilkinsem obdrželi Nobelovu cenu, teprve vyšlo najevo, že G a C se skutečně párují jako v jejich původní struktuře, ovšem s tím rozdílem, že jsou spojeny třemi můstky, a ne dvěma (obr. 2). Tvoří-li ale A–T Hoogsteenovy (HG) páry a G–C zase Watsonovy- Crickovy (WC), jak mohou existovat v jedné dvoušroubovici? Vždyť v HG párech jsou báze k sobě o hodně blíže! Pochybnosti tedy přetrvávaly. Navíc se brzy ukázalo, že páry HG mezi G a C mohou existovat také, jen C musí přijmout jeden proton navíc (obr. 2).
Rozhodnutí mohl přinést jen krystal skutečné dvoušroubovice. Tohoto cíle dosáhl Alexander Rich se spolupracovníky roku 1979, kdy publikovali první krystal krátké sekvence šesti párů bází, v níž se střídaly C a G [3]. Strukturu tvořily páry WC, dvoušroubovice však byla levotočivá. Dnes tuto strukturu nazýváme Z-DNA, neboť spojnice fosforových atomů v ní tvoří typický „cikcak“ obrazec (obr. 1C). Teprve roku 1980 získali H. R. Drew, R. E. Dickerson a spolupracovníci [4] krystalovou strukturu sekvence dvanácti párů bází, tedy delší než jedna otočka dvoušroubovice, která definitivně potvrdila model pravotočivé struktury s páry WC uvnitř (obr. 1B). Ale ani Hoogsteenovy páry neřekly poslední slovo. Vyskytují se v komplexech DNA s některými proteiny nebo s antibiotiky a rovněž jistý typ DNA‑polymerázy využívá Hoogsteenova párování při replikaci jak intaktní, tak poškozené DNA. Nedávno byly páry HG objeveny v malém množství i ve volné DNA. Zdá se, že jde o minoritní, ale nepochybně existující alternativní párování, jehož význam zřejmě ještě není doceněn.
Polymorfní a dynamická
Drewův a Dickersonův krystal nastolil snad ještě více otázek, než kolik jich zodpověděl. Jeho sekvence je CGCGAATTCGCG, tedy páry A–T jsou uprostřed a G–C na koncích. Všimněme si nejdříve, že vlákna cukrfosfátové páteře nejsou po obou stranách dvoušroubovice stejně vzdálená – na jedné straně jsou k sobě blíže než na druhé (je to dobře vidět i u párů bází na obr. 2). Tvoří tak na struktuře dva žlábky: širší (zvaný velký) a užší (malý). Na obr. 1B je uprostřed struktury k divákovi otočený malý žlábek, na koncích velký. Kdybychom molekulou otáčeli kolem osy dvoušroubovice, zjistili bychom zajímavou věc: žlábky nemají podél celé struktury stejnou šířku, konkrétně malý žlábek je ve střední části (AATT) užší než na koncích (CGCG). Šířka žlábku tedy závisí na sekvenci bází.
To však není vše. Také prostorové uspořádání bází a jejich párů závisí na sekvenci – vidíme například, že páry ve střední části jsou téměř kolmé k ose, kdežto na koncích jsou skloněné. Otočení páru kolem osy vzhledem k předchozímu (říkáme mu twist) je sekvencí zvlášť silně modulováno, i když průměrná hodnota přes všechny sekvence odpovídá 10,6 páru na jednu otočku, tedy hodnotě nepříliš vzdálené původnímu Watsonovu a Crickovu odhadu.
Ani zde se ale nemůžeme zastavit. Zatím totiž pozorujeme jen jednu určitou, statickou strukturu v krystalu. V živém organismu však molekula DNA koná tepelný pohyb a její prostorové uspořádání se stále trochu mění. Mohli bychom si myslet, že atomy prostě jen kmitají kolem svých rovnovážných poloh a nic zajímavého se neděje. Opak je však pravdou. Metoda nukleární magnetické rezonance (NMR) ukázala, že každý úsek cukrfosfátové páteře spojující dvě báze přechází rychle tam a zpět mezi dvěma konformačními stavy, které značíme BI a BII. Zastoupení těchto stavů opět závisí na sekvenci. Navíc v každém z nich je trochu jiná i prostorová konfigurace bází. Vidíme tedy, že dvoušroubovice DNA není zdaleka tak statická a stejnorodá, jak by se mohlo zdát. Jde o dynamickou, polymorfní strukturu, jejíž konformační vlastnosti jsou ovlivněny sekvencí bází, které ji tvoří.
Každý (protein) podle svého gusta

Není však tato provázanost sekvence a prostorové struktury, kterou tak jasně ukazují různé experimentální metody, vlastně jen informační šum? Vždyť příroda si možná vystačí s dvoušroubovicí a párováním bází a nestará se o to, jaký konkrétní tvar určitá sekvence má. Tak to ale vůbec není. DNA totiž není jen archivem genetické informace. Přepis genů do proteinů (exprese) musí být také regulován. Klíčovou roli zde hrají tzv. transkripční faktory – proteiny, které se vážou na specifické sekvence DNA. Jak poznají, kam se navázat? Existují dva hlavní mechanismy. Proteiny jednak mohou rozpoznávat chemické skupiny ve velkém i malém žlábku a tvořit s nimi vodíkové můstky. Mohou ale také rozpoznat sekvenčně závislou strukturu, včetně populací stavů BI a BII, a vybrat si tu, která nejlépe odpovídá jejich tvaru. Proteiny si mohou strukturu DNA také přizpůsobit, zdeformovat ji. Energie nutná k deformování DNA závisí na typu deformace, např. natočení párů bází do žlábků je mnohem snazší než jejich natočení směrem k páteři. Nepřekvapí, že energie deformace závisí také na sekvenci bází – některé sekvence se deformují snáze než jiné. To vše proteiny využívají a obvykle kombinují několik zmíněných postupů.
Stanovení souvislosti mezi sekvencí bází, tvarem a mechanickou tuhostí DNA je tedy klíčové pro pochopení vazby transkripčních faktorů i dalších molekul na dvoušroubovici. Přes značné úsilí, zahrnující nejméně tři poslední desetiletí, není tento problém uspokojivě vyřešen. Situaci komplikuje už to, že velmi podobné sekvence mohou mít velmi různé mechanické vlastnosti. Tento typ informace také není snadno přístupný experimentu – zde mohou velmi pomoci počítačové simulace. A navíc je složitá i sama povaha problému. Sekvenční závislost totiž není lokální. To například znamená, že tvar a mechanická tuhost dvojice po sobě následujících párů nezávisí jen na sekvenci bází v těchto párech, ale i na sousedních párech a v některých případech i na párech vzdálenějších.
Biologové se o vazbu transkripčních faktorů na DNA velmi zajímají. Rádi by věděli, který z nich se váže na kterou sekvenci a jak pevně (s jakou afinitou). To by jim umožnilo lépe porozumět regulaci genové exprese, která zahrnuje velmi složité sítě navzájem provázaných interakcí i úseky DNA často velmi vzdálené od samotného regulovaného genu. Nástroje pro predikci vazby transkripčních faktorů samozřejmě existují, nově jsou často založeny na metodách strojového učení. Většinou pracují pouze se sekvencí. Jejich predikční schopnost se však dramaticky zvýší, přidáme-li k sekvenci DNA ještě údaje o jejích mechanických vlastnostech. Tyto vlastnosti tak tvoří vedle samotné sekvence další vrstvu informace, kterou DNA nese.
Epigenetické modifikace
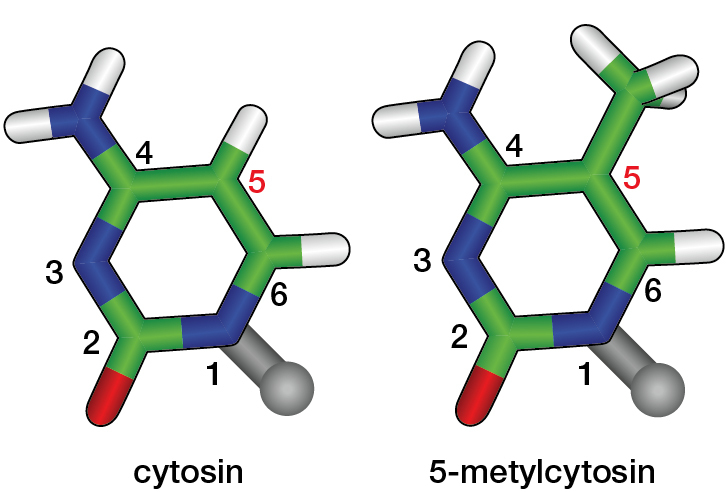
Existuje však ještě další informační vrstva. Báze totiž mohou být chemicky modifikovány přidáním metylové skupiny. V eukaryotech se může metylovat pouze cytosin (obr. 3), u obratlovců jen cytosin následovaný v sekvenci guaninem. Protože se tím nezmění genetická informace, která je v sekvenci zakódována, mluvíme nikoli o genetické, ale o epigenetické změně.
Metylovaný cytosin mC se páruje s G právě tak jako nemetylovaný. Co se tedy změní? Metylaci DNA lze chápat jako svého druhu značkování genetického textu, které udává, kdy a kde se má informace přečíst. Obecně lze říci, že metylace těch sekvencí, které regulují expresi genů, jako jsou promotory, tuto expresi potlačuje. To obstarávají proteiny, které rozeznávají metylované sekvence a spouštějí další procesy, vedoucí k tvorbě kompaktnějšího chromatinu, a tím k omezení transkripce. Přítomnost či nepřítomnost metylace je ovšem také regulována. Existují proteiny, které metylovou skupinu přidávají, jiné, které ji odstraňují, a ještě jiné, které doplňují metylaci na nově vznikající vlákno DNA při replikaci. Metylace zajišťuje stabilní expresi genů specifických pro daný typ buněk, souvisí se stárnutím (mluvíme o „metylových hodinách“) a s různými nemocemi. Příznivé vnější vlivy, jako zdravá výživa a tělesné cvičení, ale mohou ovlivnit epigenetický stav k lepšímu.
O biologickém významu metylace DNA tu jistě napíšou povolanější, nás zde zajímá jen ze strukturního hlediska. Metylová skupina v první řadě představuje velikou hydrofobní „kouli“ trčící do velkého žlábku, což, jak se zdá, rozpoznávají proteiny při navázání na metylovanou sekvenci. V poslední době se rýsuje shoda i na dalších strukturních účincích. Pokud však jde o účinek metylace na mechanickou tuhost, výsledky si zatím zcela protiřečí – není jasné, zda metylace změkčuje DNA, nebo ji vyztužuje. Efekt pravděpodobně závisí na okolních párech, ale jak, to zatím nevíme. Nedávno publikované výsledky přitom ukazují těsnou souvislost mezi těmito sousedními páry a afinitou některých proteinů. Konkrétně jde o protein s názvem DNMT1, který doplňuje metylovou skupinu při replikaci, a proteiny DNMT3A a DNMT3B, které metylují DNA úplně nanovo. Například DNMT3A a DNMT3B silně preferují T v páru o dva předcházejícím CG, kdežto DNMT1 na téže pozici nemůže vystát C, a určité preference byly pozorovány i na opačné straně CG. Taková modulace afinity sekvencí může překvapovat, čekali bychom asi, že tyto proteiny se budou vázat na všechna CG stejně silně. Ale příroda je důmyslnější, než si myslíme. Tyto přirozené preference umožňují afinitu těchto proteinů regulovat, a to na více úrovních, a nepřímo tak modulovat i vazbu transkripčních faktorů.
Jak sešroubovat šroubovice
Lidé jsou hraví, ale mají i praktické sklony a leckdy jsou vedeni jak zvědavostí, tak zištnými motivy. Tyto a další pohnutky vedly ke vzniku zcela nového oboru – DNA‑nanotechnologie. Lidé se prostě pokoušejí sestavit z DNA umělé struktury a použít je k různým účelům. Ale jak na to? Vždyť DNA je jen dvoušroubovice; přivařím-li k ní další kus, mám zase jen to samé, v podstatě jednorozměrný útvar. Nanejvýš jej mohu zacyklit. Ale abychom dostali zajímavé struktury, skutečný materiál vyrobený z DNA, potřebujeme dvoušroubovici rozvětvit.
I zde nám příroda napověděla. Nadrian Seeman, zakladatel oboru DNA‑nanotechnologií, jednou vzpomínal, jak začínal svou vědeckou kariéru jako neúspěšný krystalograf („nedokázal jsem zkrystalovat nic, co by zajímalo mne nebo kohokoli jiného“), až ho napadlo studovat dynamiku rozvětvení, které vzniká jako meziprodukt při homologní rekombinaci. To je proces nastávající během meiózy, typu buněčného dělení, dávajícího vznik pohlavním buňkám. Odpovídající chromozomy si při něm vyměňují své sekvence za vzniku
Od těchto pionýrských dob v osmdesátých letech zažívají nanotechnologie DNA prudký rozvoj. Jejich produkty slouží nejrůznějším účelům od diagnostiky po dopravu léčiv v organismu na místo určení. Základem jejich úspěchu je dobře definované párování řetězců, důležitým stavebním prvkem pak molekulové rozvětvení, stabilizované vložením dvou Hollidayových spojů za sebou (obr. 4) – vzniká tak motiv dvojitého křížení (double-crossover, DX). Spojením dvoušroubovic tímto a dalšími spojovacími prvky lze sestavit složité rovinné i prostorové útvary. Struktury DNA například často fungují jako jakési lešení, na něž jsou navázány jiné molekuly v definovaných polohách, takže mohou zcela určitým způsobem interagovat. Je důležité rozumět mechanice těchto útvarů, abychom dokázali předpovědět, jak budou reagovat na vnější zatížení a odolávat tepelnému pohybu. Rýsuje se tu nový okruh problémů – strukturní mechanika na nanoškále.
Příběh pokračuje
Sedmdesát let po svém objevení je dvoušroubovice DNA1) předmětem zájmu čisté i aplikované vědy a mediální hvězdou první velikosti. Má základní roli nejen v molekulární biologii a genetice, ale také v prudce se rozvíjejícím oboru DNA-nanotechnologií. Jako každá správná hvězda si ale také uchovává své tajemství. Počáteční jasnozřivá strukturní hypotéza Watsona a Cricka byla v odborných kruzích přijata zprvu vlažně, ještě menší byl zájem médií. Jediným celostátním britským deníkem, který jí věnoval pozornost, byl News Chronicle. V článku s mnohoslibným názvem „Blíže k tajemství života“ se čtenáři mohli dočíst, že na těchto čtyřech „prvcích“ (tj. bázích) závisí rysy předávané z generace na generaci. „Nikdo ovšem netvrdí, že tato seskupení by kdy mohla být připravena uměle,“ hlásal dále článek. Pokračoval však takto: „Objevování, jak jsou tyto chemické ‚karty‘ zamíchány a párovány, zabere vědcům dalších padesát let.“ Nebude nadsázkou, vyslovíme-li podobné tvrzení i dnes. Jen to číslo bude asi vyšší.
Poděkování: Tento článek vznikl v rámci projektu specifického vysokoškolského výzkumu (projekt č. A2_FCHT_2022_046) podpořeného Interní grantovou agenturou VŠCHT.
Poznámky
1) Řeč je o dvoušroubovici jako prostorovém uspořádání. DNA jako takovou objevil a izoloval už roku 1871 Friedrich Miescher (Vesmír 100, 312, 2021/5).
Literatura
[1] Watson J. D., Crick F. H. C.: A structure for Deoxyribose Nucleic Acid. Nature 171, 737–738, 1953, DOI: 10.1038/171737a0.
[2] Franklin R. E., Gosling R. G.: Molecular Configuration in Sodium Thymonucleate. Nature 171, 740–741, 1953, DOI: 10.1038/171740a0.
[3] Wang A. H.-J. et al.: Molecular structure of a left-handed double helical DNA fragment at atomic resolution. Nature 282, 680–686, 1979.
[4] Drew H. R. et al.: Structure of a B-DNA dodecamer: Conformation and dynamics. Proc. Natl. Acad. Sci. USA 78, 2179–2183, 1981.
Ke stažení
 článek ve formátu pdf [642,87 kB]
článek ve formátu pdf [642,87 kB]
O autorech
Filip Lankaš
Eva Matoušková
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
