









Dnešní mikroskopie v biomedicíně
 V minulých stoletích se světelné mikroskopy v biologii používaly převážně pro pozorování a popis buněk či jejich výrazných struktur. Rozvoj fluorescenční mikroskopie umožnil lokalizovat v biologických systémech i jednotlivé makromolekuly. Izolace zeleně fluoreskujícího proteinu (GFP – green fluorescent protein) z medúzy, příprava jeho barevných variant (YFP – žlutá, CFP – modrozelená ad.) a objevy dalších fluoreskujících proteinů (od modrých po červené) v mořských korálech společně s rozmachem konfokální mikroskopie (Vesmír 83, 586, 2004/10) v uplynulém desetiletí posunuly hranice poznání v buněčné biologii a rozšířily možnosti světelného mikroskopu. Mikro-skopy dnes neslouží jen k prostému popisu buněčných organel, ale pomáhají odhalovat pohyb i interakce jednotlivých molekul, především proteinů, přímo v buňkách, a tím přispívají k pochopení jejich biologických funkcí.
V minulých stoletích se světelné mikroskopy v biologii používaly převážně pro pozorování a popis buněk či jejich výrazných struktur. Rozvoj fluorescenční mikroskopie umožnil lokalizovat v biologických systémech i jednotlivé makromolekuly. Izolace zeleně fluoreskujícího proteinu (GFP – green fluorescent protein) z medúzy, příprava jeho barevných variant (YFP – žlutá, CFP – modrozelená ad.) a objevy dalších fluoreskujících proteinů (od modrých po červené) v mořských korálech společně s rozmachem konfokální mikroskopie (Vesmír 83, 586, 2004/10) v uplynulém desetiletí posunuly hranice poznání v buněčné biologii a rozšířily možnosti světelného mikroskopu. Mikro-skopy dnes neslouží jen k prostému popisu buněčných organel, ale pomáhají odhalovat pohyb i interakce jednotlivých molekul, především proteinů, přímo v buňkách, a tím přispívají k pochopení jejich biologických funkcí.
 Protein, který chceme studovat, nejdříve přímo v buňce geneticky „vybavíme“ fluorescenčním přídavkem, např. již zmíněným GFP 1) (chceme-li sledovat více proteinů současně, použijeme pro každý typ proteinu jinou barevnou variantu přídavku). Abychom mohli úspěšně popsat jeho dynamické chování, stačí navodit stav, při němž je pouze určitá část molekul tohoto proteinu fluorescenčně aktivní. Toho se dosahuje dvěma, v principu opačnými přístupy: cíleným „vybělením“ (fotodegradací) nebo fotoaktivací fluorescence.
Protein, který chceme studovat, nejdříve přímo v buňce geneticky „vybavíme“ fluorescenčním přídavkem, např. již zmíněným GFP 1) (chceme-li sledovat více proteinů současně, použijeme pro každý typ proteinu jinou barevnou variantu přídavku). Abychom mohli úspěšně popsat jeho dynamické chování, stačí navodit stav, při němž je pouze určitá část molekul tohoto proteinu fluorescenčně aktivní. Toho se dosahuje dvěma, v principu opačnými přístupy: cíleným „vybělením“ (fotodegradací) nebo fotoaktivací fluorescence.
- Vybělení fluorescence. 2) Zvolenou oblast buňky nejprve osvítíme intenzivním laserovým pulzem s vlnovou délkou odpovídající excitační vlnové délce GFP. Tím „označené“ molekuly proteinu (tj. vybavené fluorescen-čním přídavkem), které se právě nacházely v dané oblasti, ztratí svou schopnost fluoreskovat. Molekuly, které se nacházely mimo tuto oblast, schopnost fluoreskovat neztratily a postupně pronikají do sledované oblasti. Intenzitu i rychlost těchto pohybů lze pozorovat dvěma rozdílnými způsoby:
- jako opětovný nárůst fluorescence ve „vyběleném“ objemu (obrázek 1), 3)
- jako úbytek fluorescence mimo něj. 4)
- Fotoaktivace fluorescence. Příkladem je fotoaktivovatelná varianta GFP (PA-GFP). Ta je v buňkách syntetizována ve stavu, ve kterém nevykazuje žádnou fluorescenci v oblasti viditelného světla. Jestliže ji však osvítíme intenzivním zářením o krátké vlnové délce (400–420 nm), struktura molekuly PA-GFP se změní, a tím se aktivuje fluorescence v oblasti zeleného světla, obdobná jako u klasického GFP. Tato metoda tak umožňuje „označit“ molekuly, které se během fotoaktivace nacházejí ve vybrané oblasti (obrázek 4), a je tak velmi vhodná pro pozorování pohybu molekul mezi buňkami v tkáních či mezi jednotlivými strukturami v rámci jedné buňky.
 Oba popsané přístupy pak vypovídají nejen o tom, kde se sledované proteiny v buňce vyskytují a jak rychle se pohybují, ale přinášejí i odpovědi na otázky „Jak dlouho trvá přepis genu?“, „Jak rychle po skončení reakce je daný enzym znovu použit?“ ap.
Oba popsané přístupy pak vypovídají nejen o tom, kde se sledované proteiny v buňce vyskytují a jak rychle se pohybují, ale přinášejí i odpovědi na otázky „Jak dlouho trvá přepis genu?“, „Jak rychle po skončení reakce je daný enzym znovu použit?“ ap.
Kromě pohybu a lokalizace jednotlivých proteinů je důležité vědět, zda spolu sledované proteiny interagují, popřípadě kde a jak intenzivně. Technika umožňující sledovat to přímo v buňce je založena na nezářivém přenosu excitační energie (FRET – Fluorescence Resonance Energy Transfer), který popsal T. Förster v polovině 20. století. V principu jde o přenos energie mezi dvěma vhodnými fluorochromy nacházejícími se v příhodné orientaci a vzdálenosti. Pokud se molekuly fluorochromů CFP a YFP k sobě dostatečně přiblíží (typická vzdálenost pro FRET je 5 nm) a vhodně se vůči sobě natočí, je pravděpodobné, že po excitaci molekuly CFP tato molekula získanou energii nezářivě předá blízké molekule YFP, takže místo modrozelené fluorescence pozorujeme fluorescenci žlutou (obrázek 5, obrázek 6). Máme-li tedy buňku, která produkuje rekombinantní formy dvou zvolených proteinů kombinovaných s CFP, resp. YFP a tyto proteiny spolu v buňce interagují, může se stát, že budou obě základní podmínky pro nezářivý přenos excitační energie během interakce splněny a k měřitelnému přenosu energie z donoru na akceptor dojde. Přenos excitační energie představuje vhodný nástroj k detekci mezimolekulárních interakcí v buňkách. Lze tak například zjistit, kde se v buňce vytvářejí makromolekulární komplexy, které právě studujeme (obrázek 6).
 Metody světelné mikroskopie nám poskytují prostředek ke studiu uspořádání a dynamiky buněčných struktur, nicméně jejich rozlišení je příliš omezující („jen“ v řádu desetin mikrometrů). Zvýšit jej lze jinými mikroskopickými přístupy. Z nich jsou nejrozšířenější metody elektronové mikroskopie („vidí“ v řádu desetin nanometrů, tedy tisíckrát více než optické). Zmíníme se o třech, které mají velké uplatnění v biomedicínském výzkumu: o počítačové tomografii, korelaci souběžné světelné a elektronové mikroskopie a nativní elektronové kryomikroskopii.
Metody světelné mikroskopie nám poskytují prostředek ke studiu uspořádání a dynamiky buněčných struktur, nicméně jejich rozlišení je příliš omezující („jen“ v řádu desetin mikrometrů). Zvýšit jej lze jinými mikroskopickými přístupy. Z nich jsou nejrozšířenější metody elektronové mikroskopie („vidí“ v řádu desetin nanometrů, tedy tisíckrát více než optické). Zmíníme se o třech, které mají velké uplatnění v biomedicínském výzkumu: o počítačové tomografii, korelaci souběžné světelné a elektronové mikroskopie a nativní elektronové kryomikroskopii.
Počítačová tomografie v elektronové mikroskopii umožňuje ze sady snímků pořízených pod různými úhly pohledu rekonstruo-vat prostorový obraz vzorku (princip je popsán v Mikroskopii dnes, Vesmír 83, M9, 2004/3). Existuje množství vědeckých problémů, pro jejichž řešení je počítačová tomografie optimální nástroj. Jako příklad uvádíme studium organizace replikačních domén v jádře lidské buňky (obrázek 3), jejichž velikost a uspořádání jsou vhodné pro tomografickou analýzu. V České republice zatím bohužel moderní elektronový mikroskop s automatickou tomografickou výbavou postrádáme.
 Jiným nepochybně perspektivním přístupem je srovnávací (korelativní) světelná a elektronová mikroskopie, která studuje jediný jev pomocí současného použití světelné a elektronové mikroskopie. Zatímco světelná mikroskopie dovoluje pozorovat živé buňky, a tedy sledovat např. časový průběh studovaného jevu, elektronová mikroskopie díky vysoké rozlišovací schopnosti odhaluje jemné detaily, které by jinak zůstaly skryty. Světelněmikroskopický obrázek se následně porovnává s elektronmikroskopickým snímkem téže oblasti buňky a studovaná struktura se detailně popíše i z hlediska jejího uspořádání. V některých případech tento přístup nejen zpřesní světelněmikroskopická pozorování, ale pomůže odhalit i chybné závěry založené pouze na výsledcích světelné mikroskopie. Omezená rozlišovací schopnost fluorescenčního mikroskopu např. nedovolí jednoznačně určit, ve které části jadérka probíhá transkripce ribozomálních genů (obrázek 7). Matematické odstranění nezaostřené části fluorescenčního signálu (dekonvoluce) vede v tomto případě dokonce k chybnému umístění aktivních ribozomálních genů do fibrilárních center jadérka. Z elektronmikroskopického snímku je však zřejmé, že transkripčně aktivní geny jsou soustředěny výlučně v denzních fibrilárních komponentách. Vysoké prostorové rozlišení elektronové mikroskopie je v tomto případě nenahraditelné.
Jiným nepochybně perspektivním přístupem je srovnávací (korelativní) světelná a elektronová mikroskopie, která studuje jediný jev pomocí současného použití světelné a elektronové mikroskopie. Zatímco světelná mikroskopie dovoluje pozorovat živé buňky, a tedy sledovat např. časový průběh studovaného jevu, elektronová mikroskopie díky vysoké rozlišovací schopnosti odhaluje jemné detaily, které by jinak zůstaly skryty. Světelněmikroskopický obrázek se následně porovnává s elektronmikroskopickým snímkem téže oblasti buňky a studovaná struktura se detailně popíše i z hlediska jejího uspořádání. V některých případech tento přístup nejen zpřesní světelněmikroskopická pozorování, ale pomůže odhalit i chybné závěry založené pouze na výsledcích světelné mikroskopie. Omezená rozlišovací schopnost fluorescenčního mikroskopu např. nedovolí jednoznačně určit, ve které části jadérka probíhá transkripce ribozomálních genů (obrázek 7). Matematické odstranění nezaostřené části fluorescenčního signálu (dekonvoluce) vede v tomto případě dokonce k chybnému umístění aktivních ribozomálních genů do fibrilárních center jadérka. Z elektronmikroskopického snímku je však zřejmé, že transkripčně aktivní geny jsou soustředěny výlučně v denzních fibrilárních komponentách. Vysoké prostorové rozlišení elektronové mikroskopie je v tomto případě nenahraditelné.
Elektronová kryomikroskopie je zatím jediná metoda elektronové mikroskopie, která dokáže zobrazit biologický preparát v jeho nativním (přirozeném) stavu. Při použití klasických metod (Vesmír 83, 146, 2004/3) se musí biologický vzorek nejdříve vhodně fixovat, popřípadě šetrně zbavit vody (dehydratovat). To jsou však pomalé procesy, které poskytují buněčným strukturám dostatek času, aby se přeorganizovaly nebo změnily svá uspořádání. 5)
Elektronová kryomikroskopie umožňuje vodu v biologickém preparátu zachovat. Při teplotě okolo –180 °C je parciální tlak nasycených vodních par tak malý, že se voda nevypařuje ani v hodnotách vakua elektronového mikroskopu (resp. vypařuje se, ale velmi málo). Současně se veškeré chemické reakce zcela zastaví. Kdyby bylo možné biologický vzorek „zmrazit“, a potom jej při dostatečně nízké teplotě pozorovat v elektronovém mikroskopu, zůstal by hydratovaný, tedy ve svém přirozeném stavu (zůstala by zachována jeho původní jemná struktura). Jakkoli to zní jednoduše, pro úspěšné uplatnění tohoto přístupu bylo třeba řešit řadu zásadních problémů. Při obyčejném zmrazení biologického mate-riálu vznikají krystaly ledu, což vytváří mnoho artefaktů, od přerozdělení rozpustných složek až po mechanická poškození. Dosáhne-li však rychlost ochlazování zhruba milion stupňů za sekundu, vzniká nekrystalické pevné skupenství vody, amorfní led, který nemá na strukturu vzorku vliv. Tento proces se nazývá vitrifikace a její použití na biologický materiál bylo klíčovým krokem pro zavedení metod elektronové kryomikroskopie v biologii. Při vitrifikaci se vzorek zchladí (a tedy fixuje) během několika desítek mikrosekund, což mimo jiné umožní zavést tzv. časově rozlišenou elektronovou mikroskopii. Můžeme si představit, že studovanou biologickou reakci spustíme a vzorky budeme připravovat vždy po uplynutí určitého časového intervalu. Časové rozlišení se bude pohybovat v řádech jednotek až desítek sekund, ale stále to je o několik řádů rychleji než při přípravě vzorku standardní metodou.
Amorfní led je bohužel nestabilní struktura, která při překročení mezní (devitrifikační) teploty přechází do formy krystalické. Proto se po vitrifikaci vzorku musí všechny operace provádět při teplotě nižší než devitrifikační (–130 °C) a elektronový mikroskop musí být přizpůsoben pro práci za těchto teplot. Dnes jsou nezbytná kryovybavení k elektronovým mikroskopům běžně dostupná i v České republice a některá pracoviště u nás jsou jimi již vybavena.
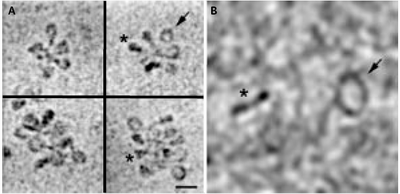
 Existují dvě základní techniky elektronové kryomikroskopie. Mikroskopie tenké vrstvy a mikroskopie nativních hydratovaných ultratenkých řezů. Mikroskopie tenké vitrifikované vrstvy se používá především ke studiu izolovaných proteinových a nukleoproteinových komplexů a částic až do velikosti asi 200 nm. Umožňuje dosáhnout subnanometrového rozlišení při zachování zcela nativního stavu biologického komplexu (obrázek 8). Tato procedura nedovoluje dodatečné kontrastování těžkými kovy, a výsledný kontrast snímků je proto velice slabý. Naštěstí pokrok v zobrazovací kvalitě elektronových mikroskopů, zejména konstrukce vysokokontrastních objektivových čoček a vysoce koherentních zdrojů elektronů, umožňuje využít fázový kontrast. Tím lze bez dodatečného kontrastování zviditelnit i takové struktury, jako jsou DNA nebo chromatinové vlákno (obrázek 2). Navíc skutečnost, že je preparát zachován ve svém původním prostorovém uspořádání, umožňuje jeho věrnou rekonstrukci. Byly to právě výsledky elektronové kryomikroskopie (obrázek 2A), které vedly k zásadní změně pohledu na uspořádání 30nm chromatinového vlákna a vyústily v zavedení „cikcak“ modelu. 6)
Existují dvě základní techniky elektronové kryomikroskopie. Mikroskopie tenké vrstvy a mikroskopie nativních hydratovaných ultratenkých řezů. Mikroskopie tenké vitrifikované vrstvy se používá především ke studiu izolovaných proteinových a nukleoproteinových komplexů a částic až do velikosti asi 200 nm. Umožňuje dosáhnout subnanometrového rozlišení při zachování zcela nativního stavu biologického komplexu (obrázek 8). Tato procedura nedovoluje dodatečné kontrastování těžkými kovy, a výsledný kontrast snímků je proto velice slabý. Naštěstí pokrok v zobrazovací kvalitě elektronových mikroskopů, zejména konstrukce vysokokontrastních objektivových čoček a vysoce koherentních zdrojů elektronů, umožňuje využít fázový kontrast. Tím lze bez dodatečného kontrastování zviditelnit i takové struktury, jako jsou DNA nebo chromatinové vlákno (obrázek 2). Navíc skutečnost, že je preparát zachován ve svém původním prostorovém uspořádání, umožňuje jeho věrnou rekonstrukci. Byly to právě výsledky elektronové kryomikroskopie (obrázek 2A), které vedly k zásadní změně pohledu na uspořádání 30nm chromatinového vlákna a vyústily v zavedení „cikcak“ modelu. 6)
 Pro pozorování nativních preparátů tkání nebo buněk se používá elektronová kryomikroskopie nativních hydratovaných řezů. Jak napovídá název techniky, jde o přípravu ultratenkých (~50 nm) řezů vitrifikovaného biologického materiálu. Metoda zahrnuje množství technologicky velice náročných postupů, od samotné vitrifikace „objemného“ vzorku přes ultratenké krájení a operace s řezy při velmi nízkých teplotách po vlastní pozorování v elektronovém mikroskopu. Přestože se tato velmi složitá metoda vyvíjí již více než dvacet let, zvládají ji pouze nejmodernější laboratoře.
Pro pozorování nativních preparátů tkání nebo buněk se používá elektronová kryomikroskopie nativních hydratovaných řezů. Jak napovídá název techniky, jde o přípravu ultratenkých (~50 nm) řezů vitrifikovaného biologického materiálu. Metoda zahrnuje množství technologicky velice náročných postupů, od samotné vitrifikace „objemného“ vzorku přes ultratenké krájení a operace s řezy při velmi nízkých teplotách po vlastní pozorování v elektronovém mikroskopu. Přestože se tato velmi složitá metoda vyvíjí již více než dvacet let, zvládají ji pouze nejmodernější laboratoře.
 Po přípravě ultratenkých řezů nebo tenké vitrifikované vrstvy následuje bez jakýchkoli dodatečných operací pozorování v elektronovém mikroskopu. I tady nastává řada obtíží, neboť vzorek je nesmírně citlivý na elektronový svazek. Kromě jiného zvyšuje jeho interakce s elektrony lokální teplotu a to může vést k následné krystalizaci vody. Vyhledávání vhodné oblasti vzorku i vlastní snímkování se proto musí provádět při minimální intenzitě svazku elektronů a typická celková dávka by neměla přesáhnout 1000 elektronů na čtvereční nanometr (tj. stokrát až tisíckrát méně než u klasického preparátu). To značně zvyšuje nároky na citlivost snímání signálu. Mikroskopy jsou proto v poslední době vybavovány citlivými CCD-kamerami, jejichž zavedení dokonce umožnilo realizovat elektronovou kryotomografii. Vůbec nejobtížnější je pravděpodobně interpretace získaných snímků. Za dobu padesáti let si biologové zvykli na vysoce kontrastní obrázky buněk a buněčných struktur a nebylo nikdy pochyb, kde například je v buněčném jádře heterochromatin a kde euchromatin. Na nekontrastovaných řezech však tento rozdíl mizí a musí se hledat rozdíly jiné, například texturální (zrnitost, vláknité uspořádání ap.). 7) Naproti tomu lze na nativních hydratovaných řezech pozorovat detaily, které jsou v klasickém preparátu překryty kontrastními barvivy, a hlavně jsou odstraněny všechny artefakty, které vznikají při přípravě vzorku standardními technikami. Je nesporné, že přínos této techniky odpovídá úsilí a prostředkům věnovaným na její povídá úsilí a prostředkům věnovaným na její vývoj. Některé vědecké výsledky nešlo získat jiným způsobem, např. údaje o uspořádání nukleozomů v tekutých krystalických fázích.
Po přípravě ultratenkých řezů nebo tenké vitrifikované vrstvy následuje bez jakýchkoli dodatečných operací pozorování v elektronovém mikroskopu. I tady nastává řada obtíží, neboť vzorek je nesmírně citlivý na elektronový svazek. Kromě jiného zvyšuje jeho interakce s elektrony lokální teplotu a to může vést k následné krystalizaci vody. Vyhledávání vhodné oblasti vzorku i vlastní snímkování se proto musí provádět při minimální intenzitě svazku elektronů a typická celková dávka by neměla přesáhnout 1000 elektronů na čtvereční nanometr (tj. stokrát až tisíckrát méně než u klasického preparátu). To značně zvyšuje nároky na citlivost snímání signálu. Mikroskopy jsou proto v poslední době vybavovány citlivými CCD-kamerami, jejichž zavedení dokonce umožnilo realizovat elektronovou kryotomografii. Vůbec nejobtížnější je pravděpodobně interpretace získaných snímků. Za dobu padesáti let si biologové zvykli na vysoce kontrastní obrázky buněk a buněčných struktur a nebylo nikdy pochyb, kde například je v buněčném jádře heterochromatin a kde euchromatin. Na nekontrastovaných řezech však tento rozdíl mizí a musí se hledat rozdíly jiné, například texturální (zrnitost, vláknité uspořádání ap.). 7) Naproti tomu lze na nativních hydratovaných řezech pozorovat detaily, které jsou v klasickém preparátu překryty kontrastními barvivy, a hlavně jsou odstraněny všechny artefakty, které vznikají při přípravě vzorku standardními technikami. Je nesporné, že přínos této techniky odpovídá úsilí a prostředkům věnovaným na její povídá úsilí a prostředkům věnovaným na její vývoj. Některé vědecké výsledky nešlo získat jiným způsobem, např. údaje o uspořádání nukleozomů v tekutých krystalických fázích.
Využití mikroskopů a nových mikroskopických přístupů v buněčné biologii prodělalo v posledních patnácti letech bouřlivý rozvoj, který stále pokračuje. Zásadním způsobem pomáhá hlouběji porozumět buněčným funkcím, ať již v základním biologickém výzkumu či v medicíně. Česká věda jej nemůže opomíjet, chce-li hrát důstojnou roli na mezinárodní vědecké scéně. Nutnou (avšak nikoliv postačující) podmínkou pro to však je, že se odpovídající přístrojové vybavení stane inventářem našich laboratoří.
Poznámky
OPTICKÁ PINZETA
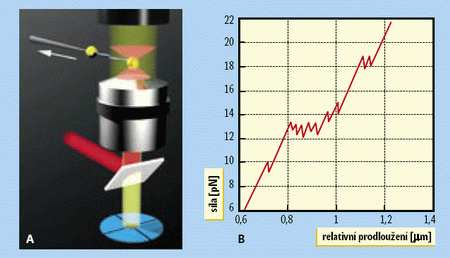 Aby naše informace o pohybu a interakcích sledovaných molekul byly úplnější, bylo by dobré změřit síly a energie nezbytné k vytvoření makromolekulárních komplexů nebo k přeskupení struktur, jejichž součástí jsou sledované bílkoviny. Jedním z moderních přístrojů, který umožňuje tyto velmi malé síly (a tedy nepřímo i energie) měřit, je optická pinzeta. Její princip objevil Arthur Ashkin koncem sedmdesátých let minulého století. V čem spočívá? Několikamikronový průhledný objekt, který se nachází v silném světelném paprsku s určitým rozdělením intenzity, bude vždy vtahován do místa s největší intenzitou. Jestliže tedy někde v prostoru vytvoříme bod, kde bude intenzita světla výrazně vyšší než v blízkém okolí, průhledný mikroobjekt bude do tohoto bodu přitažen a nebude z něj moci uniknout, dokud jej nějaká vnější síla z této „optické pasti“ nevytáhne. Vytvořit takový bod je možné pomocí zaostření intenzivního laserového svazku objektivem světelného mikroskopu s velkou numerickou aperturou (obr. A).
Aby naše informace o pohybu a interakcích sledovaných molekul byly úplnější, bylo by dobré změřit síly a energie nezbytné k vytvoření makromolekulárních komplexů nebo k přeskupení struktur, jejichž součástí jsou sledované bílkoviny. Jedním z moderních přístrojů, který umožňuje tyto velmi malé síly (a tedy nepřímo i energie) měřit, je optická pinzeta. Její princip objevil Arthur Ashkin koncem sedmdesátých let minulého století. V čem spočívá? Několikamikronový průhledný objekt, který se nachází v silném světelném paprsku s určitým rozdělením intenzity, bude vždy vtahován do místa s největší intenzitou. Jestliže tedy někde v prostoru vytvoříme bod, kde bude intenzita světla výrazně vyšší než v blízkém okolí, průhledný mikroobjekt bude do tohoto bodu přitažen a nebude z něj moci uniknout, dokud jej nějaká vnější síla z této „optické pasti“ nevytáhne. Vytvořit takový bod je možné pomocí zaostření intenzivního laserového svazku objektivem světelného mikroskopu s velkou numerickou aperturou (obr. A).
 GFP – JEDINEČNÁ SVÍTÍCÍ BÍLKOVINA
GFP – JEDINEČNÁ SVÍTÍCÍ BÍLKOVINA
Molekula zeleného fluorescenčního proteinu (GFP), izolovaná z mořské medúzy pohárovky (Aequorea victoria), má podobu soudku o průměru 3 a výšce 4 nm, v němž je uzavřen unikátní obsah: skupina tří aminokyselin (SYG – Serinu, tYrosinu a Glycinu), která při osvícení modrým světlem zeleně fluoreskuje. Je hodně jiných bílkovin, které ve svém řetězci mají stejnou sekvenci aminokyselin, a přesto nesvítí. V GFP „světlonoš uzraje“, až když se po dokončení proteosyntézy polypeptidový řetězec sbalí do podoby zmíněného soudku. Téměř uprostřed, dokonale ochráněna od okolních vlivů, se skupina SYG přemění do cyklické podoby, oxiduje a začne fluoreskovat.
Na rozdíl od ostatních fluoreskujících bílkovin, které jsou citlivé na světlo až po připojení nějaké exotické molekuly, je molekula GFP schopna provést tuto kosmetickou změnu sama na sobě, modifikací svých vlastních aminokyselin. Pro její využití ve vědě to má obrovský význam – protože nepotřebuje žádný specifický enzym ze „své“ medúzy, lze GFP připravit v libovolném organizmu. Oba konce GFP molekuly zůstávají navíc vně soudku a jsou tedy volně přístupné. Na libovolný z nich lze proto další peptidovou vazbou připojit jiné bílkoviny, a to již na genové úrovni. Stejně jako při syntéze samotného GFP, po vytvoření fúzního proteinu z GFP a studované bílkoviny se soudek GFP sám sbalí a připraví pánům vědcům k laskavému použití. Sledování míst výskytu studované bílkoviny je pak triviální – stačí si na ni posvítit modrým světlem, které budí fluorescenci GFP.
Ke stažení
 článek v pdf souboru [449,45 kB]
článek v pdf souboru [449,45 kB]
O autorech
Jan Malínský
David Staněk
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
