









Retinová kyselina, kuřecí a muší křídlo, počítačový ježek...
| 5. 2. 1995Vznik složitých orgánů a tkání z nediferencované zárodečné masy buněk patří mezi děje, které dnes vzbuzují pozornost biologů. Jde v podstatě o problém zrušení symetrie: prostor, ve kterém se zárodek či jeho část vyvíjí, se stává nehomogenním, strukturuje se, dostává vlastnosti morfogenetického pole (viz Vesmír 72, 276, 1993/5). V morfogenetickém poli mohou buňky určit svou pozici v prostoru i čase a podle toho se diferencují a vstupují do nadbuněčných vztahů. Primární struktura morfogenetického pole bývá vnucena vnějšími faktory (viz např. vývoj vajíčka octomilky, Vesmír 72, 553, 1993/10), i když jeho dynamika je posléze na vnějších faktorech autonomní. Jedním ze způsobů, jak vytvořit morfogenetické pole, je difuze morfogenu z omezené oblasti do okolí. Vznikne tím morfogenetické pole definované koncentrací morfogenu. Buňky vnímají koncentraci morfogenu ve svém okolí a podle ní se dovědí o své poloze v poli a nastaví odpovídající diferenciační program svůj i svého potomstva (obr. 1b).
Končetina obratlovců se vyvíjí z tzv. končetinového pupenu, jak je na příkladu kuřecího křídla schematicky znázorněno na obr. 1c. Už v tomto stadiu, kdy ještě nelze rozeznat žádné náznaky příští morfologie, je pupenu vtištěno schéma budoucí končetiny. Jak je vtisk proveden a jak si tkáň původní vtištění pamatuje?
Tkáň pupenu reaguje na signál z malého okrsku buněk v jeho zadní části; tento okrsek dostal jméno zóna polarizační aktivity (ZPA). Transplantací nadbytečných ZPA do různých míst pupenu lze vypěstovat končetiny s variabilním počtem prstů (obr. 1d – f). Výsledky v šedesátých letech interpretoval Wolpert tak, že ze ZPA se do prostředí vylučuje nějaký morfogen. Roku 1987 se zdálo, že hypotéza morfogenu byla potvrzena: bylo dokázáno, že v ZPA se tvoří kyselina trans-retinová (RA), která difunduje do okolní tkáně a vytváří se gradient její koncentrace (obr. 1b). Navíc, stejných výsledků jako s transplantací nadbytečné ZPA se dalo dosáhnout přikládáním tamponů namočených v RA na různá místa pupenu.
Euforie kolem retinové kyseliny nevydržela dlouho: záhy se ukázalo, že jako ZPA mohou fungovat i buňky, které byly nějakou dobu inkubovány v přítomnosti retinové kyseliny, pak zbaveny všech jejích stop a transplantovány do pupenu. Protože takto opracované buňky samy retinovou kyselinu nesyntetizovaly, bylo jasné, že retinová kyselina není primární materiální podstatou morfogenetického pole. (Vyvolává v ZPA pouze syntézu jiného morfogenu a její přítomnost je potřebná jen k definici okrsku ZPA.)

I když povaha morfogenu tak zůstávala záhadou, v devadesátých letech bylo objasněno, jakým způsobem je tvar morfogenetického pole natrvalo vtištěn buňkám pupenu a jejich potomstvu. Tímto vtiskem je exprese některých ze spektra tzv. homeotických genů (obr. 2). Jednou vyvolaný soubor produktů homeotických genů (jedna sada u členovců, čtyři u čelistnatých obratlovců) funguje jako identifikační karta každé buňky. Podle tohoto homeotického kódu se pak buňky a jejich potomci vtělují do různých struktur. Takto jsou definovány např. tělní segmenty členovců i obratlovců, diferenciace končetin, rozložení různých druhů peří na těle ptáků, ale i tvar těla nezmara. Homeotický kód končetiny obratlovců, tedy i ptačího křídla, je určen minimálně dvěma soubory homeotických genů: řada HOX D určuje rozložení prstů, řada HOX A rozložení proximodistální části, tj. ramena, předloktí atd.
A co ten ježek? Jedna z mnoha mutací popsaných u octomilky se projevuje tak, že embryo a larva jsou bohatě ochlupené a připomínají ježka (angl. hedgehog - hh). Ukázalo se, že produktem genu hhje protein, který se zásadním způsobem účastní určování polarity tělních segmentů, a to tak, že je vylučován do prostředí a 
 signalizuje buňkám, umístěným víc dopředu, jak se mají diferencovat. V nepřítomnosti proteinu hh zůstávají segmenty symetrické, nemají rozlišenu přední a zadní část. Obdobným způsobem se účastní vytváření asymetrie v zárodcích (tzv. imaginálních terčích) hmyzích končetin a křídel: místo exprese hh opět definuje zadní část a současně indukuje tvorbu struktur předních (obr. 3). Exprese genu v přední části terče způsobí zdvojení ne nepodobné situaci vyvolané u ptačího křídla transplantací ZPA či aplikací kyseliny retinové.
signalizuje buňkám, umístěným víc dopředu, jak se mají diferencovat. V nepřítomnosti proteinu hh zůstávají segmenty symetrické, nemají rozlišenu přední a zadní část. Obdobným způsobem se účastní vytváření asymetrie v zárodcích (tzv. imaginálních terčích) hmyzích končetin a křídel: místo exprese hh opět definuje zadní část a současně indukuje tvorbu struktur předních (obr. 3). Exprese genu v přední části terče způsobí zdvojení ne nepodobné situaci vyvolané u ptačího křídla transplantací ZPA či aplikací kyseliny retinové.
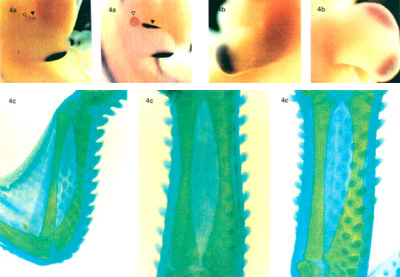
Vraťme se k ptačímu křídlu. C. Tabin se spolupracovníky, ve snaze nalézt morfogen vylučovaný ze ZPA, identifikoval protein, který byl syntetizován buňkami ZPA a vylučován do prostředí. Byla-li jeho syntéza vyvolána na nepatřičných místech pupenu, vyvolával stejné změny morfogeneze jako výše popsané transplantáty ZPA, aplikace retinové kyseliny nebo transplantace buněk ovlivněných retinovou kyselinou (Riddle R. D. a spol.: Cell 75, 1401, 1993). Záhy se ukázalo, že tento protein je homologem proteinu Hedgehog z octomilky, dostal proto jméno Sonic hedgehog (Shh) podle populární postavičky počítačových her. (Příznačné je, že článek vyšel na Silvestra.) Na barevné příloze (obr. 4a – c) lze vidět expresi genu Shh, homeotického genu D13 a vývoj křídla při normální a experimentální ontogenezi pupenu.
Ve stejném čísle časopisu Cell jsou hned dva další články o roli proteinu Shh při diferenciaci nervové soustavy v zárodku savců a v embryogenezi ryb, v obou případech opět v roli induktora 
asymetrie v dané struktuře (obr. 6a – b). Lze se domnívat, že všechny tři případy, popsané na různých modelech, budou záhy prohlášeny za univerzální. Exprese Shh v končetinovém pupenu savců byla už také prokázána (obr. 4d a obr. 6a).
Interpretace funkce proteinu Shh (a jeho analogu Hh u octomilky) je dvojí. Může fungovat jako klasický morfogen a vytvářet koncentrační gradient s dlouhým dosahem, nebo zůstává na místě produkce a bude ovlivňovat jen buňky bezprostředně přiléhající k ZPA. Takto ovlivněné buňky vytvoří jiný signál, kterým ovlivní další vrstvu buněk atd., až se touto tichou poštou strukturuje celý prostor pupenu. V každém případě je vytvořená asymetrie dále fixována - např. vzorcem exprese homeotických genů. Tento vzorec pak uchovávají všichni potomci ovlivněných buněk pupenu.
Vypadá to, že při rozčlenění tělního plánu používají všichni živočichové několik univerzálních nástrojů. Takovými nástroji mohou být malé signální molekuly typu retinové kyseliny nebo růstových faktorů, nebo proteiny - produkty exprese regulačních genů (homeotické geny, Hh resp. Shh a mnoho dalších). Tyto nástroje jsou natolik univerzální, že obvykle nejsou potíže s rozpoznáním homologických genů (resp. z nich odvozených proteinů) u různých skupin živočichů. Navíc tyto geny přenesené z obratlovce do octomilky a naopak jsou v novém prostředí obvykle schopny zhostit se funkce svého přirozeného homologa.
Opět tedy máme možný morfogen. Příznačná je reakce prof. Wolperta: "Nádhera... Měli bychom se z toho radovat, dokud je nám to dovoleno. Už jsme jednou zažili podobnou euforii u retinové kyseliny. Vždy se nakonec ukáže, že věci jsou komplikovanější, než jsme si mysleli."
Obrázky

O autorovi
Anton Markoš
Doporučujeme

Ideologie v mapách, mapy v rukách ideologů 

Pátranie po pôvode SARS-CoV-2 pokračuje
