










O egoizmu všeho živého
Přijmeme-li jednou darwinizmus jako možné vysvětlení světa kolem, zjistíme s překvapením, že svět takto nahlížený krásně funguje. Organizmy mají maximalizovat svůj reprodukční úspěch – a skutečně to dělají, když takové činění předpokládáme. Ptáček zpívá, ale už nezpívá jen tak pronic zanic – zpívá buď proto, aby přilákal samici na svůj zpěv, nebo aby obhájil teritorium před jinými samci a přilákal na svoje teritorium samici. Ježek nezpívá a napohled vlastně nic nedělá, ale protože víme, že kdyby něco dělal, bude tím maximalizovat svůj reprodukční úspěch, žádné ježkovo chování, kdybychom je objevili, nás nepřekvapí.
Sobecký organizmus a sobecký „gen“
Jsou ovšem taková různá chování organizmů, která nelze jasně převést do jazyka individuálního egoizmu. Například altruizmus. Včelí dělnice, která se nikdy nemnoží, a samice kulíka, která na sebe připoutává pozornost predátora, působí dojmem egoistů poněkud nedokonalých a zřejmě neúspěšných. Po mnoha letech osvěty snad už všichni tuší, v čem je zde jádro pudla: Ti altruisté nejsou altruističtí jen tak naslepo, nýbrž selektivně vůči svým blízkým příbuzným, s nimiž sdílejí nějaký podíl společných genů. Obětovat se – v jakémkoliv smyslu – pro svého bratra nemá smysl, neboť tento bratr je (statisticky) pouze poloviční kopií mne samotného (a to ještě pouze za předpokladu, že máme společného otce, což není nikterak zaručeno); obětovat se pro více než dva bratry a více než osm bratranců už se může vyplatit. Věc má ovšem háček – já jsem já, a můj egoizmus se tedy týká mé osoby, nikoliv nějakých částečných kopií mé osoby v cizích tělech; mně po nějakých bratrancích nic není, protože můj bratranec rozhodně nejsem já. Je-li tu někdo, kdo mne ponouká, abych riskoval svůj život ve prospěch více než osmi bratranců (příležitost jistě vzácná), rozhodně to nejsem já, ale někdo jiný. Kdo?
No přece ty geny, odpovídá Richard Dawkins. Geny se chtějí množit, replikovat (proto „replikátory“), a aby se jim to povedlo, postavily si mne. Protože ale stejná kopie stejného genu se s padesátiprocentní pravděpodobností vyskytuje v mém bratru (tedy pokud otec...), je tomuto genu úplně jedno, kterým tělem, zda mým či bratrovým, pronikne do další generace. Nejraději oběma, tedy dvakrát, samozřejmě, ale když to nejde, já a bratr se nelišíme. Z mého osobního pohledu je mezi mnou a jakkoliv příbuzným ne-mnou značný rozdíl, z hlediska sobeckého replikátoru často nikoliv. Egoizmus organizmů je prostě jenom součet egoizmů jednotlivých replikátorů; jedinec úspěšně maximalizující svůj reprodukční úspěch prostě jenom úspěšně maximalizuje reprodukční úspěch replikátorů, které v něm bydlí. Úspěšné egoistické replikátory jsou ty, které si dokázaly vyrobit úspěšné tělo. Dokonce i altruizmus jedince je prostě jenom součet egoizmů jednotlivých replikátorů, neboť jejich rozšíření není omezeno na jedno jediné izolované tělo: altruizmus je pokračování egoizmu jinými prostředky.
Dawkinsovská reformace darwinizmu přinesla zásadní změnu pohledu – není to nová teorie, je to nový jazyk, umožňující popisovat jediným způsobem mnoho jevů druhdy konfliktních. David Storch mluvívá o galileovském obratu, o nahlédnutí relativity či jiného vztahu, jenž právě pro svou relativitu může být popsán z několika stran. (Po letech opakování této jeho myšlenky cítím povinnost ji konečně citovat.) Ještě přesnější by ale možná byl „obrat einsteinovský“. Heliocentrizmus koneckonců geocentrické nazírání v jakémsi smyslu vyvrátil a nahradil je novým; teorie relativity klasické fyzice nic takového neprovedla, pohybujeme-li se v normálním světě. Newtonovská fyzika popisuje reálný svět lidských rozměrů dobře a dodnes, zatímco Einstein prostě jenom rozšířil fyzikální jazyk do oblastí, o nichž Newton nemluvil. Dawkins nám umožňuje mluvit i o věcech, s nimiž by klasický darwinizmus dosti ztěžka zápolil; jsou to jevy většinou mladší (tedy později objevené) než samotný dawkinizmus a dodatečně jej, řekl bych, velmi posilují.
Proti „teorii sobeckého genu“ bývá vznášeno mnoho námitek, vesměs nejapných („molekula přece nemá morálku, takže přece nemůže být sobecká, přece!“), neúplně japných (např. Gould: Pandin palec), či poněkud ideologicky zabarvených (např. Markoš: Povstávání živého tvaru). Nechce se mi teď polemizovat, snad jindy; ale zřejmé je, že si za to může Dawkins sám užíváním slova „gen“. To slovo totiž nic neznamená, respektive to, co si pod ním představují molekulární biologové, nemá konceptuálně nic společného s tím, co myslí Dawkins, s tím, čemu on sám s Hullem říká „replikátor“ a čemu Williams říká „kodexová (nebo kodikální) doména“. Především by nám mělo být jasné, že to, co nás zajímá, je informace a že DNA je prostě médium, kterým se informace šíří; DNA není zpráva, ale papír, na němž je nějaká zpráva (eventuálně) napsána. Gen je informace, gen není věc. Dnes to zní divně a kacířsky, ale pojem „gen“ tak, jak byl zaveden, znamenal „vlohu“, tj. informaci pro vytváření vlastnosti. Buňka, ale taky chromozom a to, čemu molekulární biologové říkají „gen“, správněji „cistron“ (tedy v podstatě úsek DNA kódující informaci o stavbě RNA a eventuálně proteinu), jsou pak pouze interaktory, tedy hmotné nosiče přímo či nepřímo určené informací, svou vlastní strukturou, svým uspořádáním přenášející tuto informaci a získávající energii pro přenos informace. Rozdíl mezi „genotypem“ a „fenotypem“ je rozdíl mezi zprávou na jedné straně a dopisem, obálkou, poštovní schránkou a ministrem pošt, tedy útvary, jimiž je zpráva přenášena, na straně druhé. Ani papír, ani inkoust nejsou „zpráva“, nýbrž její nosiče; DNA není genotyp, nýbrž fenotyp, ale molekulární. Nic, co můžeme centrifugovat, purifikovat, vážit a sekvencovat, není a už jaksi z definice nemůže být „genotyp“. Gen v našem smyslu je prostě to něco, co se množí klonálně, a to nezávisle na jiných takových „něcech“; to něco, co přežívá rozklad jedince, meiózu, oplození a vznik nového jedince, aniž by se to samo změnilo; a to něco, co se vyskytuje v několika variantách různě úspěšných, pokud jde o šíření. Vůbec nezáleží na chemické podstatě toho něčeho; obvykle to bývá kousek DNA, ale pokud to bude kus membrány nebo cytoskeletu, nepřestane to proto být evolučním genem, tj. replikátorem.
Mimotělní fenotyp aneb o neexistenci organizmů
Psi poznají lidi čichem, ale nerozeznají jednovaječná dvojčata, která sdílejí stejný genotyp. Psi cítí naše geny, anebo naopak: genetická informace člověka určuje stavbu DNA (a je určována stavbou DNA), stavbu RNA, stavbu proteinů, stavbu buněk a orgánů, chování člověka a chování psů, mimo jiné. Chování psů k člověku je součástí jeho i jejich fenotypu. Schopnost psa vyčenichat zločince je kriminalisticky závažná, ale pro evoluci člověka nehrála žádnou roli (spíše pro evoluci psa – takoví bloodhoundi vznikli do značné míry proto, aby hledali uprchlé otroky). Schopnost vrtejší larvy žijící v blešivci přinutit blešivce, aby plaval k hladině, kde ho sežere kachna, v níž larva vrtejše dospěje a dokončí svůj vývoj, je podstatným faktorem v evoluci všech zúčastněných. Součástí fenotypu rehka je jeho zobák i jeho hnízdo, součástí fenotypu kukačky je její zobák i její způsob vyhledávání rehčích hnízd, ale také způsob reakce rehka na krásné velké mládě, které se mu nějak udělalo. Krmí-li rehek kukaččí mládě, nezískává tím pražádnou výhodu, naopak, a nelze si tedy představit selekční proces, jenž by přiměl rehčí geny determinovat chování takto podivné. Kukaččí geny, které vyrobí mládě splňující všechny rehčí sny o správném potomkovi, jsou evolučně úspěšné – a chování rehka je tedy fenotypová extenze kukaččích, nikoliv rehčích genů („extended phenotype“ podle Dawkinse). Genotypu odpovídá fenotyp – ano, ale tento fenotyp se může projevovat na těle jedince, v němž geny sídlí (zobák), úplně mimo tělo (hnízdo), a dokonce i na cizím těle (rehkovo poblouznění). Sobectví informace tedy není omezeno na médium, v němž je zapsána; sobectví genu přesahuje fyzickou existenci interaktoru, který byl genem sestrojen.
 Jakási mutace způsobí, že myši mají porušený orgán rovnováhy ve vnitřním uchu, takže se melou kolem dokola. Říkáme jim „tančící myši“, čímž jaksi naznačujeme, že kaskáda změn vyvolaná mutací v nějakém genu skončila poruchou chování. Ale kdybychom tyto myši zkoumali mrtvé (což v mnoha případech biologové vskutku dělají, neboť naprostou většinu známých druhů vlastně nikdy nikdo neviděl zaživa), byla by tatáž mutace označena nějak jako „abnormální vnitřní ucho“. A samozřejmě, zjistíme-li, že tančící myši jsou velmi ohroženy dravostí jiných zvířat neboli predací (což by, v případě, že by nežily v teráriu, vskutku byly), můžeme mluvit o mutaci způsobující „predační zranitelnost“. Každý gen má několik fenotypových úrovní, jimiž může být popsán, včetně úrovní „mimotělního“ (extendovaného) fenotypu. Je tedy nakonec rozumné mluvit o genech pro bobří přehradní jezera – jestliže se vyvinulo bobří jezero jako adaptace (k něčemu), musela být na počátku nějaká variabilita ve velikosti, tvaru, hloubce, umístění apod. přehrad, přesněji řečeno variabilita v genech tyto parametry jezer ovlivňujících. Hloubka jezera je fenotyp bobra; pravda, fenotyp zprostředkovaný bobřím chováním stejně jako bobří chování jsou zprostředkovány neuroanatomií a neurofyziologií, které samy jsou dány biochemií. Z druhé strany: geny přežívají pomocí svých fenotypových účinků, a je prostě fakt, že většina fenotypových projevů, ale ne všechny, se usazuje na těle organizmu. Organizmus je pro replikátory věc obvyklá, ale nikoliv nezbytná – jak ještě uvidíme.
Jakási mutace způsobí, že myši mají porušený orgán rovnováhy ve vnitřním uchu, takže se melou kolem dokola. Říkáme jim „tančící myši“, čímž jaksi naznačujeme, že kaskáda změn vyvolaná mutací v nějakém genu skončila poruchou chování. Ale kdybychom tyto myši zkoumali mrtvé (což v mnoha případech biologové vskutku dělají, neboť naprostou většinu známých druhů vlastně nikdy nikdo neviděl zaživa), byla by tatáž mutace označena nějak jako „abnormální vnitřní ucho“. A samozřejmě, zjistíme-li, že tančící myši jsou velmi ohroženy dravostí jiných zvířat neboli predací (což by, v případě, že by nežily v teráriu, vskutku byly), můžeme mluvit o mutaci způsobující „predační zranitelnost“. Každý gen má několik fenotypových úrovní, jimiž může být popsán, včetně úrovní „mimotělního“ (extendovaného) fenotypu. Je tedy nakonec rozumné mluvit o genech pro bobří přehradní jezera – jestliže se vyvinulo bobří jezero jako adaptace (k něčemu), musela být na počátku nějaká variabilita ve velikosti, tvaru, hloubce, umístění apod. přehrad, přesněji řečeno variabilita v genech tyto parametry jezer ovlivňujících. Hloubka jezera je fenotyp bobra; pravda, fenotyp zprostředkovaný bobřím chováním stejně jako bobří chování jsou zprostředkovány neuroanatomií a neurofyziologií, které samy jsou dány biochemií. Z druhé strany: geny přežívají pomocí svých fenotypových účinků, a je prostě fakt, že většina fenotypových projevů, ale ne všechny, se usazuje na těle organizmu. Organizmus je pro replikátory věc obvyklá, ale nikoliv nezbytná – jak ještě uvidíme.
Vůbec se musíme rozhodnout, co si budeme představovat pod pojmem „jedinec“: buď konkrétní genotyp, poskládaný ze zásoby genů, kterou má populace k dispozici, anebo konkrétní fyziologicky nezávislou materiální entitu. Obvykle je to jedno a totéž, třeba člověk – ale už jednovaječná dvojčata jsou jeden genotypový jedinec ve dvou fyziologických jedincích. V botanice se užívají pojmy genet a ramet a není důvodu neaplikovat je i na jiné organizmy. Perloočky se zhusta množí partenogeneticky, tedy klonálně; z jedné zygoty vznikne během letní sezony v rybníku klon milionů těl (rametů), roztroušených v planktonu a obvykle se nikdy nepotkávajících, nicméně tvořících jednoho genetického jedince, jeden genet. Úspěšnost tohoto genetu je dána kolektivní schopností všech rametů přenést geny do další pohlavní generace. Naši dva ministerští náměstkové Svobodové (Josef S., bývalý náměstek ministra kultury, a jeho jednovaječný bratr Cyril S., bývalý náměstek ministrů spravedlnosti a zahraničních věcí, nyní ministr vnitra) jsou dva ramety jednoho genetu, jehož fenotypem je bytí náměstkem ministra něčeho.
Renegáti
Různé replikátory se šíří různými způsoby a jejich strategie šíření jsou to, co determinuje jejich vlastnosti i jejich osud. Pro další výklad bude užitečné vnímat organizmus, v němž replikátory sedí, jako jejich hostitele; zkusme se na (sobecký) replikátor podívat jako na parazita.
Vezměme si třeba motolici, tedy organizmus, který parazituje v jiném organizmu. Má nejen vlastnosti umožňující jí, aby svému hostiteli odebírala živiny, ale také vlastnosti, jimiž se svým hostitelem manipuluje. Tak třeba motolice druhu Dicrocoelium dendriticum se (prý!) zavrtá do mozku mravence a jaksi ho programuje, aby vylezl na vrchol stébla a čekal tam, dokud ho nesežere ovce, neboť je v zájmu motolice dostat se do ovčího střeva; v zájmu mravence to zjevně není. Tohle je důležité – nejde o situaci, kdy nakažený mravenec jen tak pomalu leze, a proto je snáze sežrán (což by byl pouhý vedlejší účinek nakaženosti), nýbrž o situaci, kdy nakažený mravenec dělá věci naprosto nesmyslné. Nejde to tedy vysvětlit pouhou oslabeností, pouhou nemocí; manipulační vysvětlení je zde patrně lepší.
Cílem parazita je dostat se v příští generaci opět do vhodného hostitele a jednou z vhodných strategií je prostě hostitele nikdy neopouštět. Dokáže-li parazitovat v zárodečných buňkách, obvykle ve vajíčkách, dostane se přes ně do zygoty, a tedy i do další generace hostitelů. Jakmile se parazit naučí přenášet se touto zkratkou, tedy vertikálně-maternálně, z matky na potomky, vyplatí se mu, aby samicím hostitele, v nichž žije, co nejmíň škodil. Parazit smí svému hostiteli kodit pouze tehdy, má-li možnost se šířit horizontálně, má-li možnost poškozeného, třeba i umírajícího hostitele opustit. Koneckonců, opouštění hostitele často způsobuje hostitelovu likvidaci. Navíc parazit šířící se vajíčky musí usilovat o maximalizaci množství vajíček, které jeho hostitelka produkuje, čili nesmí jí vůbec škodit. Znamená to, že parazit omezený na maternální přenos má stejné zájmy jako kterýkoliv hostitelčin gen: nadělat co nejvíc životaschopných potomků své hostitelky.
Ale pozor – parazit se dostane do zygoty, nicméně polovina parazitů se dostane do samčí zygoty, z níž další producent vajíček nevznikne; polovina parazitů tedy končí ve slepé uličce, protože stěží mohou ovlivnit, která spermie oplodní právě to jejich vajíčko. Pro ně pak může být výhodné svého nevýhodného hostitele zlikvidovat a jít dál. Někdy to dělají. Mikrosporidie v komárech zůstávají avirulentní, když zjistí, že jsou v samici, a naopak zabijí svého hostitele a infikují ekundární hostitele (buchanky), když zjistí, že jsou v samci. Jiní paraziti v savcích, přenášení placentou nebo mlékem do potomků, zůstávají v klidu, jsou-li v samici, a ožijí a množí se, pouze když tato samice zabřezne; naopak, jsou-li v samci, množí se okamžitě a pokoušejí se dostat z hostitele ven. Ještě lepší je přeměnit samce, v němž jsem se při své smůle octl, v samici; i to se děje (viz např. cytoplazmatické bakterie v suchozemských korýších). Anebo přinutit samici, v níž bydlím, aby se nemnožila sexuálně (a neprodukovala tedy potomky obojího pohlaví), nýbrž partenogeneticky, tedy aby produkovala pouze své dokonalé kopie, tj. dcery. Anebo aspoň zabít samce, když už v něm jsem; tím samozřejmě nezachráním sebe, neboť uniknout nemohu, ale mohu tím pomoci svým vlastním genetickým kopiím (svým „sestrám“) obývajícím samice (hostitelovy sestry) někde okolo. Pokud třeba samci a samice soupeří o nějaký zdroj, zvýhodní chcípnutí samce okolní samice, a tím jsou zvýhodněni i paraziti v těchto samicích. Hranice mezi parazitickým organizmem a egoistickým replikátorem hostitelského organizmu se nám poněkud stírá. Koneckonců, vnitrobuněčný parazit nikdy neopouštějící buňku a šířící se pouze spolu s jejím množením je vlastně organela; mitochondrie a chloroplasty určitě vznikly ze symbiotických (parazitických?) bakterií.
V přenosu genetické informace zkrátka existují zájmy a konflikty zájmů. Dva genetické elementy mají stejné zájmy, pokud sdílejí stejný způsob přenosu. Parazit i hostitel sdílejí přání, aby hostitel žil, ale mohou se lišit v přání, jak dlouho má hostitel žít. Dle hostitele je dobré žít aspoň do rozmnožení sebe sama, dle parazita musí hostitel žít tak dlouho, aby parazit stihl svůj vývoj v něm a aby ho mohl opustit; pouze vertikálně se šířící parazit má stejný zájem na délce života hostitele jako hostitel sám, a to jenom tehdy, nachází-li se v hostiteli správného pohlaví, obvykle v samici.
Také gen sedící na chromozomu se může chovat jako parazit. Tak například gen segregation distorter (sd) u octomilky se vyskytuje ve dvou formách, agresivní a neagresivní, a ovlivňuje jiný gen, řídící výrobu spermií, přičemž řízený gen opět existuje ve dvou formách, náchylné a otužilé. Agresivní sd produkuje toxin, jímž likviduje náchylnou formu ovládaného genu, ale ne formu otužilou. (Je to samozřejmě složitější, ale nám teď nejde o realitu, nýbrž o model.) Oba geny sedí na stejném chromozomu velmi blízko u sebe, takže se šíří skoro výhradně spolu. Existují tedy čtyři možné kombinace: agresivní-náchylný, agresivní-otužilý, neagresivní-náchylný, neagresivní-otužilý. První kombinace, pokud někdy vznikne, spáchá sebevraždu, takže se jí nemusíme zabývat; poslední dvě kombinace obsahují neagresivní formu genu sd, takže se nic neděje. Agresivní gen sd může sedět na jednom chromozomu pouze spolu s otužilou formou ovládaného genu, takže s formou náchylnou se může potkat pouze v oplozeném vajíčku, kam náchylná forma přišla z té druhé zárodečné buňky. Máme tedy zygotu, a pak i jedince nové generace, v jehož buňkách se sešel agresivní-otužilý chromozom s chromozomem neagresivním-náchylným. Agresivní sd nemůže poškodit svůj chromozom (neboť ten obsahuje otužilou verzi), ale zlikviduje ten druhý chromozom; zabrání tedy vzniku spermií, v nichž by sídlila konkurenční, neagresivní verze. Výledkem této bitvy tedy je, že jakmile se agresivní forma genu sd potká s neagresivním konkurentem, zabrání mu přejít do další generace: nadělá spermií sice míň, ale všechny budou obsahovat agresivní typ. A protože spermií je vždycky nadbytek, vyplatí se to. Tento gen se zkrátka chová jako parazit, ačkoliv skoro určitě nevznikl z nějakého cizího symbionta.
Je to prostě normální gen, který se vymkl kontrole; je to gen, který se vzbouřil. Takovým genům se říká různě, někdy dosti poeticky (psanci, renegáti), jindy méně poeticky (sobecké genetické elementy, sebeprosazující elementy, ultrasobecké geny). Důležité je, že ostatní geny samozřejmě nejsou existencí těchto odpadlíků nijak nadšeny. Neagresivní verze genu sd nehyne se špatně vyrobenou spermií sama; agresivní sd zabíjí i všechny ostatní geny, které se náhodou připletly do stejné vznikající spermie jako jeho oběť. Cílem ostatních genů je tedy odpadlíka zlikvidovat, anebo mu pomoci. Jakmile totiž pobije své konkurenty, nebude mít koho pobíjet a stane se neškodným. (Sociální analogie této situace nechť si laskavý čtenář dosadí sám.) Po genomu jsou tedy navíc roztroušeny také geny, které rvačkám ultrasobeckých elementů brání, a to jak supresory, které jejich šíření omezují, tak enhancery, které je podporují.
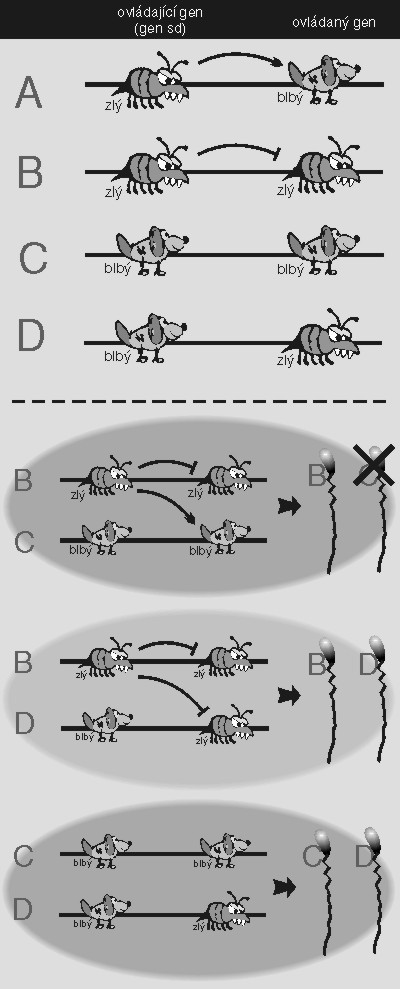 Lze říci, že ultrasobecký element usilující o své namnožení má několik možných strategií, jak to učinit. Může se množit uvnitř buňky, pokud se nějak vymkne mitotické kontrole sčítající chromozomy při buněčném dělení (třeba tak, že se rozlézá po chromozomu, nebo tak, že sedí v cytoplazmě). Může přeskakovat z buňky do buňky, tedy šířit se horizontálně; v tom případě promořuje populaci nezávisle na tom, jak ovlivňuje úspěšnost svých hostitelů. Chová se pak jako klasický parazit, pouze vznikl zevnitř, osamostatněním – tak patrně vznikly viry. A může se šířit spolu se svým hostitelem, a to tak, že zvyšuje jeho úspěšnost; to dělají klasické jaderné geny, které máme ve zvyku považovat za normální.
Lze říci, že ultrasobecký element usilující o své namnožení má několik možných strategií, jak to učinit. Může se množit uvnitř buňky, pokud se nějak vymkne mitotické kontrole sčítající chromozomy při buněčném dělení (třeba tak, že se rozlézá po chromozomu, nebo tak, že sedí v cytoplazmě). Může přeskakovat z buňky do buňky, tedy šířit se horizontálně; v tom případě promořuje populaci nezávisle na tom, jak ovlivňuje úspěšnost svých hostitelů. Chová se pak jako klasický parazit, pouze vznikl zevnitř, osamostatněním – tak patrně vznikly viry. A může se šířit spolu se svým hostitelem, a to tak, že zvyšuje jeho úspěšnost; to dělají klasické jaderné geny, které máme ve zvyku považovat za normální.
Není tomu tak – pomáhat svému hostiteli je jenom jedna z možností jak se množit, a to zjevně možnost nikoliv nejvýhodnější. Organizmus je kartel genů a jeho celková úspěšnost je ovlivněna především těmi nejméně úspěšnými geny, které v něm sídlí a které s genem, o jehož strategii se zajímáme, sdílejí stejný způsob přenosu. Je marné mít svaly, dobrou povahu a rozum, ocitnou-li se geny kódující tyto znamenitosti v jednom těle s geny kódujícími nedostatečnou loajalitu k vrchnosti, slepotu a sklon k černému moru. Také dialog dvou lidí se vždy vede na úrovni toho hloupějšího.
Ale ještě jedna možnost existuje: sedět v jádře na chromozomu a ovlivňovat organizmus ve svůj prospěch, bez ohledu na jeho prospěch, třeba i v jeho neprospěch. Sem patří segregation distorter a jemu podobné geny, sem ale patří i geny pracující v neprospěch jednoho pohlaví.
Válka rodičů s dětmi
Je známo, že v zygotě, a tedy i v embryu, nemusejí být aktivní obě kopie nějakého genu, mateřská a otcovská, nýbrž pouze kopie jediná, a to nenáhodně (tzv. genomový imprinting). Některé geny se zkrátka dědí jako funkční jednotky pouze po linii jednoho pohlaví. Nesmíme zapomínat, že jedinec je mozaika mateřských a otcovských genů, které nemusejí sdílet stejné zájmy. Pokud se matka páří s několika samci, má gen embrya zděděný po otci nezcela odhadnutelnou šanci, že v sousedním embryu se nachází stejná kopie tohoto genu; gen zděděný po matce má tuto šanci padesátiprocentní, gen zděděný po otci maximálně padesátiprocentní. V děloze matky zkrátka existují současně – z hlediska embrya – tři typy genů: moje embryonální geny zděděné po matce, moje embryonální geny zděděné po otci a ostatní geny v matce (a v sourozencích). Matka rozděluje živiny co nejrovnoměrněji, protože nemá žádný zájem dávat přednost jednomu embryu na úkor jiných, neboť všechna jsou jí stejně příbuzná. Embryonální geny samozřejmě chtějí získat co nejvíc živin, čímž se jejich úspěšnost zvýší – a jen málo se jejich úspěšnost sníží, když sousední embryo bude hladovět (ledaže by sousední embryo bylo opravdu sourozenec, ale to nikdo neví). Otcovské embryonální geny jsou samozřejmě sobečtější, protože nemají žádnou záruku, že v sourozencích bydlí jejich kopie a ne geny od nějakého jiného otce. Je to zkrátka stejná situace, jako když rodiče ponoukají dítky, aby se o pamlsky dělily rovným dílem (neboť z hlediska rodičů jsou všichni potomci stejně příbuzní, a tedy zaměnitelní), k čemuž dítky nejeví žádnou ochotu, protože můj sourozenec je mi sice příbuzný na padesát procent (pokud ten pán, který si říká náš společný otec...), ale já jsem sám sobě příbuzný stoprocentně, a tedy víc. Srovnáme-li fakt genomového imprintingu (nenáhodné, jednorodičovské aktivity jediné kopie genu v zygotě a embryu) s touto složitou nitroděložní strategií, vycházejí nám některé předpovědi a můžeme se podívat, budou-li splněny. Zdá se, že něco na tom bude: aktivní je otcovská kopie růstového faktoru Igf2 a mateřská kopie jeho receptoru Igf2r. Otec drží pod kontrolou gen ovládající sobecký růst embrya (na úkor sourozenců, kteří nemusejí být jeho půlce dítěte vůbec příbuzní), matka se brání kontrolou receptoru, který tento růstový faktor do buňky pustí, nebo (raději) nepustí. Nicméně nutno přiznat, že tento poněkud bojovně trockistický pohled na dělohu je dosti nový, na nevelkém materiálu založený – a s alternativními koncepcemi soutěžící. Přesto se domnívám, že je to hypotéza pěkná, a navíc podložená i dalšími podivnými jevy.
Předpokládejme tedy, že embryo se snaží urvat co nejvíc mateřských živin, a tedy látky uvolňované embryem do mateřského těla jsou tu od toho, aby manipulovaly matkou a nutily ji dodávat co nejvíc živin nikoliv sourozencům, nýbrž mně. Nezapomínejme, že embryo v lůně matky není součást matky, nýbrž jiný jedinec, a to z 50 % cizí jedinec, vetřelec. Embryo by přinejmenším rádo, aby živiny, třeba glukóza, zůstaly v matčině krvi co nejdéle a byly mu k dispozici, nikoliv aby je matka spotřebovala sama. Lze tedy očekávat, že embryo se bude snažit udržet glukózu v krvi, a tedy pracovat proti mateřskému inzulinu. Opravdu, placentární laktogen (hPL) a placentární růstový hormon (hPGH) jsou vlastně antiinzuliny; placenta totiž, kdo to nevíte, nevzniká z matky, ale z embrya, takže placentární buňky obsahují těch 50 % otcovských, tudíž cizích genů. Otcovské geny chtějí mít velkou placentu, aby matku hodně vysály, mateřské naopak; a opravdu: když jsou v zygotě dvě prvojádra, mateřské a otcovské, nahrazena dvěma otcovskými, embryo si udělá velkou placentu, a když jsou nahrazena dvěma mateřskými, placenta se zmenší. (Bohužel nikoliv bez výjimek.)
Bojovníci proti potratům by si tedy měli být vědomi, že interrupce prostě přeruší tenhle boj, asi jako když od sebe roztrhneme dva rvoucí se psy; že jasně preferujeme jednoho z protivníků, není nijak nepochopitelné, uvědomíme-li si, kdo z těch dvou platí daně. Jinak je tento boj tlumen prostě jenom tím, že embryo nemá svou matku vyhladovět k smrti, aby neumřelo, a matka nemá vyhladovět svůj plod, aby zanechala potomstvo; ti dva se potřebují, a proto uzavřeli přechodnou koalici. Maminka, tatínek a děťátko jsou něco jako Stalin, Roosevelt a Churchill v Jaltě. Ale ne víc.
Tato koncepce genetických konfliktů má, znovu opakuji, alternativy; nicméně je nejelegantnější, neboť dokáže vysvětlit jedním jazykem i detailní znalosti, dosud považované za vcelku nedůležité marginální faktíky. Elegance není důkazem pravdivosti; ale nejsme-li schopni najít pravdu, začne být elegance dosti důležitá. Kosmologové a subnukleární fyzikové to vědí: nikdy nebudeme tzv. vědět, jak vznikl vesmír, ale příběhy, které si na to téma vymýšlíme, nechť jsou jednoduché, celistvé a pěkné.
Poučení
Zdá se, že má stále menší smysl mluvit o organizmech, o jedincích, v jiném než praktickém, tedy víceméně jatečním smyslu. Organizmus je přechodný jev, který nemá žádnou unikátní evoluční strategii, neboť je v krátké době dezintegrován smrtí; i jeho genotyp záhy zanikne buď smrtí, nebo rekombinací v průběhu pohlavního množení. Množí-li se organizmus nepohlavně, není mu pak zase konce: jedincem z evolučního hlediska není perloočka, nýbrž mnoho perlooček. To, co biologové obvykle zkoumají, tedy samostatná fyziologická entita (ramet), není žádná evolučně podstatná jednotka.
Žije-li organizmus na skále jako lišejník, je prostě buď živý, anebo neživý. Žije-li organizmus v cizí buňce, může se postupně rozplývat, ztrácet – jako kočka Šklíba, ztrácející se od ocasu, až z ní zbude jenom škleb. Hranice organizmu jsou zcela nejisté, ať už zevnitř (představte si sekvenci cytoplazmatická bakterie → mitochondrie → mitochondriální gen v jádře; pokud je virus nebo segregation distorter nebo ultrasobecký cytoplazmatický gen součástí organizmu, nemá organizmus žádné svoje jednotné zájmy, pokud není jeho součástí, rozpadá se nám organizmus před očima na spoustu samostatných entit), anebo zvenčí (kukačka → chování kukačky → chování rehka ke kukačce).
Organizmus je prostě jakési časoprostorové zahuštění sobeckých zájmů různých replikátorů. Je to koalice replikátorů, které sdílejí společnou strategii přenosu do budoucnosti – koalice replikátorů se stejnými předpověďmi budoucnosti. Člověk a tasemnice nesplývají, neboť jejich očekávání nejsou zcela totožná; člověk a tasemnice jsou jaksi dva organizmy.
A teď zkusme trochu vyměnit slova: společnost je jakési časoprostorové zahuštění sobeckých zájmů různých lidí. Chovat se podle morálních pravidel, tedy konformně vůči společnosti, je zkrátka optimální většinová strategie, což je ten optimistický prvek, ale není to strategie jediná, což je ten pesimistický prvek. Lze loupit a lze neloupit; většina lidí bude vždycky neloupit a menšina bude vždycky loupit; tak jako vždy v minulosti.
To neznamená, že ti, kdož neloupí, nejsou egoisté – prostě jenom volí jinou strategii maximalizace svého zisku. Někdo zkrátka uplatňuje svůj egoizmus jako bankéř nebo advokát nebo univerzitní profesor, a někdo dělá totéž jako pán s tyčí, čekající v nočním parku na náhodné chodce. Můžeme si říkat, že bankéři, advokáti a univerzitní profesoři společnosti škodí vcelku méně a že evoluce by měla směřovat k postupné eliminaci mužů s tyčí, ale nemáme žádný důvod předpokládat, že se to někdy stane. Jedinci maximalizují svůj zisk, jak se jim to v dané situaci nejlépe hodí, a předpokládáme-li, že býti bankéřem nebo advokátem nebo univerzitním profesorem vyžaduje jakési vnitřní mohutnosti, jimiž ne každý vládne, musíme se smířit s tím, že ti ostatní maximalizují svůj osobní zisk nějak jinak někde jinde. Protože ovšem nemohou všichni členové populace postávat v nočních parcích, můžeme optimisticky předpokládat, že počet nerafinovaných zlosynů a intenzita prostě jadrného násilí bude v populaci pořád stejná; nejenže jich neubude, ono jich také nepřibude. Ani univerzitními profesory, ani muži v parku nemohou být všichni; ten dělá to a ten zas tohle; a někdo to dělat musí.
Vrátíme-li se ke genům – všechny jsou sobecké, přičemž některé vytvořily koalici genů majících stejné zájmy, to jest vytvořily organizmus; ostatní se šíří nějak jinak, bez ohledu na organizmy i proti jejich zájmům, a to jsou geny ultrasobecké. Jsou lidé sobečtí, jako já, vy, kdo to čtete, Albert Schweitzer nebo Viktor Kožený, a jsou lidé ultrasobečtí, jako například Babinský. Ultrasobečtí jedinci pracují proti našim zájmům a jest jim za to pykati; nicméně nemáme žádný důvod se vysilovat pohoršováním nad jejich špatností: jdou na to jenom jinak.
Možná to řekněme ještě jasněji: máme stejné cíle jako oni, ale jdeme na to jinak.
Ke stažení
 Článek ve formátu PDF [422,76 kB]
Článek ve formátu PDF [422,76 kB]
O autorovi
Jan Zrzavý

Doporučujeme

Ničí ozon choleru? 

Jak se člověk stává biologem
