










Akumulátorky života
 Předloni se na stránkách Vesmíru (Vesmír 75, 309, 1996/6) objevil článek prof. O. Nečase „Molekulární motory“ o proteinech – motorech proměňujících energii uvolněnou štěpením adenozintrifosfátu (ATP) na energii mechanickou. Mluví se v něm o pohybech zprostředkovaných těmito motory v součinnosti s cytoskeletem. Letošní Nobelova cena za chemii nám dává příležitost přiblížit jiný, evolučně mnohem starší druh molekulárních motorů: membránové proteinové komplexy – pumpy.
Předloni se na stránkách Vesmíru (Vesmír 75, 309, 1996/6) objevil článek prof. O. Nečase „Molekulární motory“ o proteinech – motorech proměňujících energii uvolněnou štěpením adenozintrifosfátu (ATP) na energii mechanickou. Mluví se v něm o pohybech zprostředkovaných těmito motory v součinnosti s cytoskeletem. Letošní Nobelova cena za chemii nám dává příležitost přiblížit jiný, evolučně mnohem starší druh molekulárních motorů: membránové proteinové komplexy – pumpy. 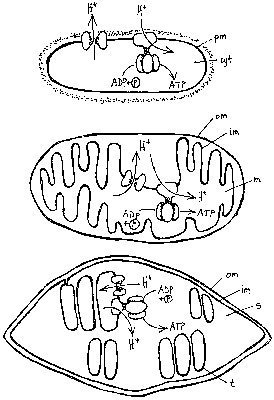 I tyto molekulární stroje štěpí ATP (proto se v jejich názvu objevuje jméno ATPáza) a využívají uvolněnou energii k nuceným přesunům látek – obvykle iontů – přes membránu. Tak se obě prostředí oddělená membránou liší v koncentraci příslušného iontu; vznikne membránová „baterie“, iontový článek schopný při svém vybití konat práci. Tento druh využitelné energie se nazývá rozdíl elektrochemických potenciálů příslušného iontu.
I tyto molekulární stroje štěpí ATP (proto se v jejich názvu objevuje jméno ATPáza) a využívají uvolněnou energii k nuceným přesunům látek – obvykle iontů – přes membránu. Tak se obě prostředí oddělená membránou liší v koncentraci příslušného iontu; vznikne membránová „baterie“, iontový článek schopný při svém vybití konat práci. Tento druh využitelné energie se nazývá rozdíl elektrochemických potenciálů příslušného iontu.
Existuje jedna velmi specializovaná pumpa, lokalizovaná do speciálního typu membrán, které se říká F0F1ATPáza. Ta může štěpit ATP a energii využívat k pumpování protonů přes membránu a vytvořit tak na ní rozdíl elektrochemických potenciálů protonů. V obvyklých případech však pracuje v opačném směru a je odpovědná za většinu ATP syntetizovaného v biosféře; pak nese název H+-ATPsyntáza. Díky tomuto zvláštnímu enzymu existují ve většině buněk dvě univerzální zásobárny energie, jež pohánějí veškeré procesy vyžadující energii. Jsou to ATP a rozdíl elektrochemických potenciálů různých iontů na membránách. Jak lze dospět k jedné nebo druhé formě, to vysvětluje rámeček na následující straně.
Laureáti
Za objasnění mechanizmu syntézy ATP enzymovým komplexem H+-ATPsyntázou získali cenu dva biochemici: Paul D. Boyer (*1918) z Kalifornské univerzity v Los Angeles a John E. Walker (*1941) z Laboratoře pro molekulární biologii v Cambridži (V. Británie). Za objev pumpy Na+K+-ATPázy, enzymu odpovědného v živočišných buňkách za elektrochemický potenciál sodných a draselných iontů na jejich povrchové (plazmatické) membráně, a za objasnění mechanizmu její funkce dostal cenu Jens C. Skou (*1918) z Århuské univerzity v Dánsku.
Za objev pumpy Na+K+-ATPázy, enzymu odpovědného v živočišných buňkách za elektrochemický potenciál sodných a draselných iontů na jejich povrchové (plazmatické) membráně, a za objasnění mechanizmu její funkce dostal cenu Jens C. Skou (*1918) z Århuské univerzity v Dánsku.
Dýchání a dobíjení buněčných „baterií“
Dýchání je vynález patrně tak starý, jako je život sám. Vyskytuje se v obou skupinách „prokaryont“ – archebakteriích a eubakteriích. Jde o geniální uspořádání: dýchací enzymy (neboli enzymy dýchacího řetězce) jsou umístěny v plazmatické membráně bakterie a přes ně „tečou“ elektrony z oxidovaných živin na vhodnou redukovatelnou látku – akceptor elektronů. Protékající elektrony odevzdávají dýchacím enzymům část své volné energie a ta je využita k pumpování protonů z cytoplazmy ven. Protože membrána je pro protony nepropustná, vznikne baterie spočívající v rozdílu elektrochemického potenciálu protonů (ΔμH+). Fotosyntetizující bakterie vynalezly k této soustavě nástavbu: při nedostatku oxidovatelného materiálu si elektrony oplývající energií připraví tím, že absorbují foton a jeho energii využijí k odebrání elektronů sloučeninám sice hojným, ale jinak energeticky jako zdroj elektronů nepoužitelným – např. vodě. S využitím energie fotonu se elektrony vynesou na vysokou energetickou hladinu, a jejich část pak opět teče systémem oxidačně-redukčních protonových pump, velmi podobných pumpám dýchacím: výsledkem procesu je rovněž ΔμH+.Protonová baterie a ATP
Protonová baterie je zdrojem energie pro mnoho buněčných funkcí. Pohání mj. bakteriální bičík (obrázek) a umožňuje (systémem tzv. kotransportu) akumulaci živin z okolí. Nemůže ale pohánět procesy, které nejsou lokalizovány přímo v membráně – např. syntézu DNA, RNA, proteinů, cukrů a dalších látek. A zde se evoluci podařil jeden z rozhodujících průlomů, když vynalezla komplex H+ATPsyntázy. Celý komplex připomíná v elektronovém mikroskopu razítko trčící z membrány (viz obrázek)). V membráně je komplex proteinů F0, který funguje jako protonový kanál, a ten je propojen s komplexem F1, vyčnívajícím do cytoplazmy. Jsou-li jednotkou F0 vpuštěny protony (po spádu svého ΔμH+), lze jejich energii zachytit a využít ji v komplexu F1 k syntéze ATP. A malá molekula ATP (obr. 2) už není vázána na membránu a může v buňce snadno difundovat tam, kde je jí třeba.Buňky eukaryontní jsou zřejmě výsledkem opakovaných symbióz eubakteriálních i archebakteriálních buněk. Jedním z pozdních počinů evoluce bylo soužití se dvěma typy eubakteriálních buněk, z nichž časem vznikly dýchací organely – mitochondrie a fotosyntetizující organely – chloroplasty. Oxidačně-redukční protonové pumpy a H+-ATPsyntáza jsou v prvním případě umístěny ve vnitřní mitochondriální membráně, a to tak, že protony jsou pumpovány z mitochondriální matrix ven a ATP následně vzniká uvnitř – v matrix. V chloroplastech jsou tyto komplexy umístěny v membráně tylakoidů – protony jsou pumpovány do tylakoidu a ATP vzniká ve stromatu chloroplastu (obr. 1).
Sekundární baterie
ATP produkovaný v mitochondriích je využíván k pohonu nejrůznějších funkcí buňky. Jednou z těchto funkcí je i pohon různých pump a tvorba sekundárních elektrochemických potenciálů iontů na membránách, které samy nemají oxidačně-redukční pumpy. V plazmatických membránách živočišných buněk je nejdůležitější pumpou Na+K+-ATPáza, udržující výrazné rozdíly v koncentracích obou iontů mezi buňkou a jejím okolím. V lidském těle je nitrobuněčná koncentrace sodných iontů asi 5 mM, ve tkáňovém moku a krevní plazmě asi 150 mM; rozložení draselných iontů je v obráceném poměru. Oba dva elektrochemické potenciály jsou – podobně jako ten primární – využívány k nejrůznějším transportům přes membránu a odpovídají také za elektrické jevy na membráně – membránový potenciál klidové buňky a jeho depolarizaci – jako způsob rychlé a účinné signalizace. Při těchto dějích, ale i spontánně, se sodnodraselná baterie vybíjí a Na+K+-ATPáza má za úkol její kapacitu obnovovat. Úkol je to náročný – klidová buňka, která zrovna „nic nedělá“, spotřebuje k pumpování sodných a draselných iontů 30 % své produkce ATP, buňky specializované k transportu (ledvinové tubuly) nebo vedení vzruchu (mozkové neurony) dokonce až 70 %.J. Skou byl u výzkumu Na+K+-ATPázy od samého počátku. Roku 1957 dokázal, že jde o pumpu poháněnou ATP, popsal její dvě podjednotky α a β a mechanizmus i stechiometrii přenosu obou iontů (obr. 4). Tím otevřel teoretickou i metodickou cestu, po které se ubíraly celé zástupy biochemiků popisujících nové a nové pumpy, jejich činnost a význam.
Syntéza ATP
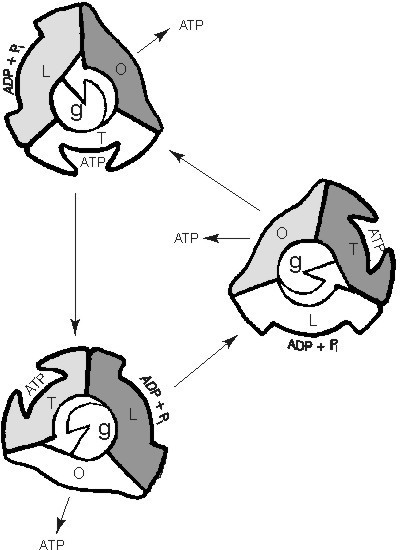
Zpět k syntéze ATP v komplexu F1. Reakce ADP s anorganickým fosfátem je silně endergonická (spotřebuje značné množství energie) a probíhá pouze v nevodném prostředí. Potřebná práce je tedy vynaložena hlavně k izolování obou reagujících molekul od okolní vody. P. D. Boyer v 60. a 70. letech nepřímo (pomocí náročných pokusů) odvodil průběh celého děje, J. E. Walker tuto hypotézu potvrdil, když připravil celou F1 v krystalickém stavu (samo o sobě úctyhodný výkon) a podrobil ji rentgenostrukturální analýze.Celý komplex F1 se skládá z 9 proteinů: α3β3γδε  . Samotná hlavička je složena ze tří dimerů αβ, ostatní podjednotky jsou ve spojovací části – stonku. ADP a fosfát, resp. ATP, se vážou na protein β. Boyer vyslovil a experimentálně podpořil hypotézu, že celá syntéza ATP probíhá ve třech taktech O, L, T. Konformace O (open) je prázdná, L (loose) váže ADP a fosfát, a T (tight) váže ATP. Současně všechny tři dimery αβ kooperují: vazba ADP a fosfátu na konformaci O znamená přechod do konformace L, přičemž sousední podjednotka přejde z L do T a dalším uvolněním molekuly ATP z T do O (obr. 3). Největším triumfem celého modelu bylo objasnění transmisního systému od komplexu F0 (tok protonů) k F1. To, co Boyer odvodil v 60. a 70. letech z dat biochemických a Walker v letech devadesátých z dat krystalografických, bylo letos potvrzeno přímo: proudem protonů se roztočí podjednotka γ a její rotace dodává energii trojtaktnímu motoru podjednotek αβ (viz rámeček 1).
. Samotná hlavička je složena ze tří dimerů αβ, ostatní podjednotky jsou ve spojovací části – stonku. ADP a fosfát, resp. ATP, se vážou na protein β. Boyer vyslovil a experimentálně podpořil hypotézu, že celá syntéza ATP probíhá ve třech taktech O, L, T. Konformace O (open) je prázdná, L (loose) váže ADP a fosfát, a T (tight) váže ATP. Současně všechny tři dimery αβ kooperují: vazba ADP a fosfátu na konformaci O znamená přechod do konformace L, přičemž sousední podjednotka přejde z L do T a dalším uvolněním molekuly ATP z T do O (obr. 3). Největším triumfem celého modelu bylo objasnění transmisního systému od komplexu F0 (tok protonů) k F1. To, co Boyer odvodil v 60. a 70. letech z dat biochemických a Walker v letech devadesátých z dat krystalografických, bylo letos potvrzeno přímo: proudem protonů se roztočí podjednotka γ a její rotace dodává energii trojtaktnímu motoru podjednotek αβ (viz rámeček 1).
ATP v učebnicích
 Největším zadostiučiněním pro vědce je situace, kdy se jeho dílo stane klasikou, učebnicovým textem, který každý považuje za samozřejmost a už se jej neobtěžuje citovat. Někdy se povědomí o tvůrci dochová aspoň v názvu (Krebsův cyklus, Mitchellova chemiosmotická teorie), jindy ani to ne. To je osud Boyera a Skoua, velikánů poválečné biochemie, a obrovského intelektuálního úsilí, které bylo jejich zásluhou uvedeno v chod. Už několik generací studentů se učí podle jejich schémat, vtlouká si do hlavy stechiometrii pumpování iontů či syntézy ATP, aniž by jim jméno Boyer nebo Skou cokoli říkalo. Jakýmsi vrtochem osudu tenkrát Nobelovu cenu nedostali, a pak „zlidověli“ natolik, že notoricky známou faktografii si už téměř nikdo nespojuje s jejími tvůrci. Nakonec měli to štěstí, že žili dost dlouho, a tak se mohou těšit z dalšího vrtochu – tentokrát švédské Královské akademie, která si na ně vzpomněla přes propast několika desetiletí...
Největším zadostiučiněním pro vědce je situace, kdy se jeho dílo stane klasikou, učebnicovým textem, který každý považuje za samozřejmost a už se jej neobtěžuje citovat. Někdy se povědomí o tvůrci dochová aspoň v názvu (Krebsův cyklus, Mitchellova chemiosmotická teorie), jindy ani to ne. To je osud Boyera a Skoua, velikánů poválečné biochemie, a obrovského intelektuálního úsilí, které bylo jejich zásluhou uvedeno v chod. Už několik generací studentů se učí podle jejich schémat, vtlouká si do hlavy stechiometrii pumpování iontů či syntézy ATP, aniž by jim jméno Boyer nebo Skou cokoli říkalo. Jakýmsi vrtochem osudu tenkrát Nobelovu cenu nedostali, a pak „zlidověli“ natolik, že notoricky známou faktografii si už téměř nikdo nespojuje s jejími tvůrci. Nakonec měli to štěstí, že žili dost dlouho, a tak se mohou těšit z dalšího vrtochu – tentokrát švédské Královské akademie, která si na ně vzpomněla přes propast několika desetiletí...
Rotace v biologii
Na úrovni makroskopické ani mikroskopické nic podobného rotačnímu motoru v živých tělech nenajdeme. Znamenalo by to fyzické oddělení rotoru od zbytku těla. A tak jsou kruhové pohyby (např. kroužení paží, ale i pohyb bičíku spermie či prvoka) řešeny složitým skládáním pohybů lineárních. Skutečnou rotaci nalezneme až na úrovni molekulární, a nanomotory mají rozměry řádově 10 nm. 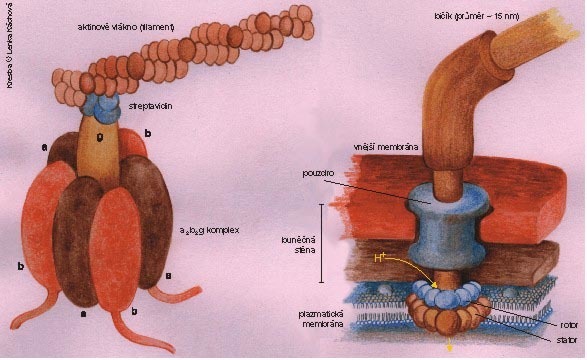 Oba dosud známé rotační motory jsou umístěny v membráně a pohání je baterie elektrochemického potenciálu protonů lokalizovaná na téže membráně. Potenciál se vybíjí tak, že protony proudí přes membránovou jednotku po svém koncentračním spádu, a při tom roztáčejí rotor.
Oba dosud známé rotační motory jsou umístěny v membráně a pohání je baterie elektrochemického potenciálu protonů lokalizovaná na téže membráně. Potenciál se vybíjí tak, že protony proudí přes membránovou jednotku po svém koncentračním spádu, a při tom roztáčejí rotor.
Obecné schéma energetických proměn v buňce
 Se zanedbatelnou výjimkou světelné pumpy halobakterií (Archaea) jsou jediným primárním zdrojem energie pro buňky reakce oxidačně-redukční, tj. tok elektronů z donoru (oxidovatelné organické nebo anorganické sloučeniny) na akceptor (redukovatelné organické nebo anorganické sloučeniny). Převážná část donorů i akceptorů elektronů je původně sama produktem endergonických (energii spotřebovávajících) oxidačně-redukčních reakcí poháněných světlem (např. organické látky jako donor, kyslík jako akceptor). Energie uvolněná tokem elektronů z donorů na akceptory je využita dvěma způsoby:
Se zanedbatelnou výjimkou světelné pumpy halobakterií (Archaea) jsou jediným primárním zdrojem energie pro buňky reakce oxidačně-redukční, tj. tok elektronů z donoru (oxidovatelné organické nebo anorganické sloučeniny) na akceptor (redukovatelné organické nebo anorganické sloučeniny). Převážná část donorů i akceptorů elektronů je původně sama produktem endergonických (energii spotřebovávajících) oxidačně-redukčních reakcí poháněných světlem (např. organické látky jako donor, kyslík jako akceptor). Energie uvolněná tokem elektronů z donorů na akceptory je využita dvěma způsoby:
- Vznikají fosfoestery organických sloučenin; transesterifikací lze fosfátovou skupinu přenést přímo na ATP. Typickým příkladem tohoto spřažení oxidačně-redukční reakce se syntézou ATP je glykolýza.
- Takzvané oxidačně-redukční pumpy použijí uvolněnou energii k přenosu protonů (někdy i jiných iontů) přes membránu. Výsledkem je rozdíl koncentrací protonů na obou stranách membrány, jehož schopnost konat práci charakterizuje forma energie nazvaná rozdíl elektrochemických potenciálů protonů. Oxidačně-redukční pumpy se vyskytují ve 4 typech membrán: v plazmatických membránách většiny archebakterií i eubakterií a ve dvou typech eukaryontních organel, jež jsou samy deriváty eubakterií ve vnitřní membráně mitochondrií a v membráně tylakoidů chloroplastů. V prvních třech případech je tento systém pump označován jako dýchací řetězec.
Rozdíl elektrochemických potenciálů protonů (druhotně i jiných iontů) na membráně je vedle ATP jednou ze dvou univerzálních baterií kdykoli dostupných k nejrůznějším formám práce. Využívá se k transportu živin, zesílení signálů, k termogenezi a u bakterií k pohonu bičíku. Prostřednictvím H+-ATPsyntetázy (F0F1) mohou obě formy energie přecházet jedna v druhou: obvyklejší směr (v chloroplastech a většině mitochondrií) je syntéza ATP, esterifikace ADP fosforečnanem. Naprostá většina ATP produkovaná v biosféře vzniká právě touto reakcí. ATP (ať už je produkováno kterýmkoli z obou systémů) pohání řadu buněčných funkcí: metabolické syntézy, pohyb, luminiscenci a také nejrůznější pumpy. Na úkor energie uvolněné štěpením ATP mohou pumpy generovat (elektro)chemické potenciály nejrůznějších látek i na membránách jiných typů, než jsou výše vyjmenované. Jedním z těchto systémů je sodnodraselná pumpa Na+K+-ATPáza na plazmatických membránách živočišných buněk. I tato forma elektrochemického potenciálu se podobně jako původní potenciál tvořený protony dá využít k práci, v tomto případě zejména k přenosu látek přes membránu a k vedení vzruchu v nervové a svalové buňce.
Ke stažení
 článek ve formátu pdf [660,81 kB]
článek ve formátu pdf [660,81 kB]
O autorovi
Anton Markoš
Doporučujeme

