









Buněčné dělení
Dělení majetku je vždycky problém, ať už po rozpadu státu či manželství. Při dělení majetku buněk jde ovšem o otázku života a smrti, protože jenom kompletně vybavená buňka může přežít a množit se. Každá buňka proto před rozdělením udělá geniální tah, který by jistě vyřešil i mnohé potíže v rozvodových síních – svůj majetek zdvojnásobí. Pak stačí, aby každá buňka obdržela půlku, a vše je v pořádku. Zatímco u některých položek na přesném rozdělení zas tolik nezáleží, mnohé statky je třeba podělit co nejpřesněji. Například pokud při rozdělení obdrží každá dceřiná buňka asi tak půlku ribozomů, jsou všichni zúčastnění zcela spokojeni. Naproti tomu s dělením dědičné informace uložené na chromozomech je zapotřebí naložit mimořádně pečlivě a přesně, protože zhruba s půlkou DNA moc štěstí neuděláme.
Replikace a koheze
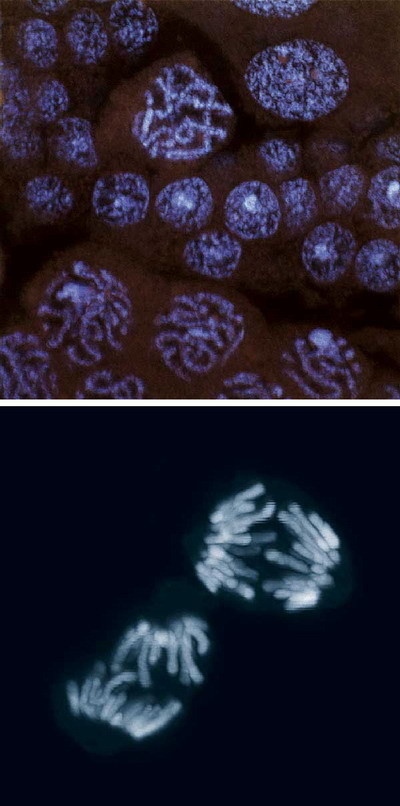
Jak je tedy zajištěno, že každá dceřiná buňka dostane kompletní genetickou výbavu? (Viz obrázek) Prvním důležitým krokem je zdvojnásobení dědičné informace uložené v chromozomech, které se uskutečňuje ve velmi složitém a přesně časovaném procesu zvaném replikaces. Během ní probíhá vedle kopírování DNA řada dalších důležitých procesů. DNA je v buňce uložena v podobě chromozomů, jež jsou kompaktním smotkem nukleových kyselin a mnoha bílkovin. Během kopírování DNA je zapotřebí chromozomy nejdříve rozbalit, aby ji bylo možné číst a opsat, a poté zase obě nové verze zabalit a připravit k rozdělení do dceřiných buněk. Kvalita celého procesu replikace je průběžně kontrolována (viz rámeček).
Během ní probíhá vedle kopírování DNA řada dalších důležitých procesů. DNA je v buňce uložena v podobě chromozomů, jež jsou kompaktním smotkem nukleových kyselin a mnoha bílkovin. Během kopírování DNA je zapotřebí chromozomy nejdříve rozbalit, aby ji bylo možné číst a opsat, a poté zase obě nové verze zabalit a připravit k rozdělení do dceřiných buněk. Kvalita celého procesu replikace je průběžně kontrolována (viz rámeček).
Výzkum v posledním desetiletí ukázal, že naprosto nezbytným krokem k spolehlivému rozdělení dědičné informace dceřiným buňkám je ustanovení koheze mezi nově replikovanými kopiemi chromozomů – tzv. sesterskými chromatidami. (Buněčné rodinné vztahy jsou poněkud zmatené – během dělení dostane každá dcera jednu sestru.) Díky tomu zůstanou dvě nové kopie chromozomu pevně spojeny. Proč je tento krok tak důležitý? Představme si, co je vlastně úkolem buňky při dělení. Jádro obsahuje několik různých chromozomů, v případě lidských buněk je jich 46. Každý chromozom dá během replikace vznik dvěma sesterským chromatidám, buňka jich tedy bude obsahovat 92. Každá dcera musí dostat zase 46 kusů, ale ne 46 jakýchkoliv chromozomů, nýbrž právě 46 různých chromozomů. Buňka však nemá žádnou možnost každou jednotlivou sesterskou chromatidu označit např. 1A, 1B, 2A, 2B, 3A, a poté přidělit všechny verze A do jedné dcery a verze B do druhé. Nejjednodušší technikou je držet sesterské chromatidy svázané těsně při sobě, nedovolit jim, aby se promíchaly, a přestřihnout pouta, teprve když je všechno dokonale připraveno k vlastnímu rozdělení buňky. Představte si to, jako když dostanete na starost desatero identických dvojčat. Vždy jedno z každé dvojice má jít do kina, druhé na plavání. Nejjednodušší je, když se dvojčata chytí za ruce, seřadí se, a pak jenom jedno jde nalevo, druhé napravo – a správné rozdělení je zaručeno. Koheze umožňuje právě ono „chycení se za ruce“.
Kohezi zajišťuje několik různých proteinů, které tvoří komplexy zvané koheziny. Hlavní osou kohezinů jsou proteiny rodiny SMC, které mají velmi charakteristickou strukturu umožňující pevné, ale pružné spojení sesterských chromatid (obrázek). Vzhledem k důležité úloze, kterou mají pro správné rozdělení chromozomů, není divu, že byly nalezeny zatím ve všech organizmech všech říší, od bakterií po člověka. Jejich důležitost potvrzuje i skutečnost, že jakákoliv mutace v kohezinech je pro buňku letální.
Kinetochor
Vytvoření kopií chromozomů a jejich propojení kohezí samozřejmě nestačí. Chromozomy je totiž nezbytné nějak dopravit na opačné strany buňky, aby po oddělení dceřiných buněk zůstala v každé buňce jedna sada chromozomů. Dopravu zajišťují mikrotubulová vlákna vytvořená speciálně pro tento účel (Vesmír 79, 438, 2000/8). Každý chromozom se připojí na jedno nebo několik vláken pomocí pozoruhodné buněčné struktury zvané kinetochor, což je velký komplex tvořený mnoha bílkovinami (u kvasinek se na jeho tvorbě podílí 60–70 různých bílkovin, u vyšších eukaryont se počet odhaduje na 80 bílkovin). Kinetochorový komplex se netvoří kdekoliv na chromozomu, ale pouze v oblasti specifické nukleotidové sekvence zvané centromera – a protože každý chromozom má jenom jednu centromeru, vytvoří se na každém také jenom jeden kinetochor (obrázek). Každý organizmus má centromeru trochu odlišnou a jako obvykle má pivní kvasinka centromeru nejmenší a nejjednodušší, zatímco centromery ostatních řádných eukaryotických organizmů jsou mnohem větší a značně složité. Funkci však mají vsechny centromery stejnou – zajistit vytvoření kinetochoru, a tím i napojení chromozomu na mikrotubuly.Co to jsou mikrotubuly a odkud se vlastně berou?
Vlákna mikrotubulů jsou nezbytnou součástí eukaryotického cytoskeletu a podílejí se na spoustě procesů, např. na pohybu buňky či na transportu různých nákladů z jedné strany buňky na druhou. Řídí je centra organizace mikrotubulů (MTOC), která jsou také mimořádně důležitá pro správnou segregaci (rozdělení) chromozomů. Jeden konec mikrotubulů, „– konec“, je v centru zakotven a nemění se. Druhý, „+ konec“, vede velmi dynamický život. Roste, zkracuje se, vyhmatává svou cestu buňkou, naráží do buněčných struktur a za vhodných okolností nakonec někde zakotví. Jedním z míst určených k zakotvení je právě kinetochor. Pokud mikrotubulové vlákno na kinetochor narazí, připojí se k němu a chromozom je připraven k transportu.Jak ale buňka rozezná, že jsou všechny kinetochory, a tedy i všechny chromozomy, připojeny správně? Dlouho nebylo jasné, jak to funguje. Dnes na tom nejsme o moc lépe, ale aspoň máme pěknou a testovatelnou (i když obtížně) hypotézu. Jak už víme, po replikaci jsou dvě nové kopie drženy pevně při sobě molekulami kohezinu. Každá kopie má centromeru, na níž se vytvořil komplex kinetochoru. Buňka je schopna kontrolními proteiny ověřit, zda je každý kinetochor připojen k mikrotubulovým vláknům. V případě, že jsou v buňce stále ještě volné, nepřipojené kinetochory, pozdrží buňka dělení až do doby, než i ty nejlínější mikrotubuly najdou v kinetochoru pevné ukotvení.
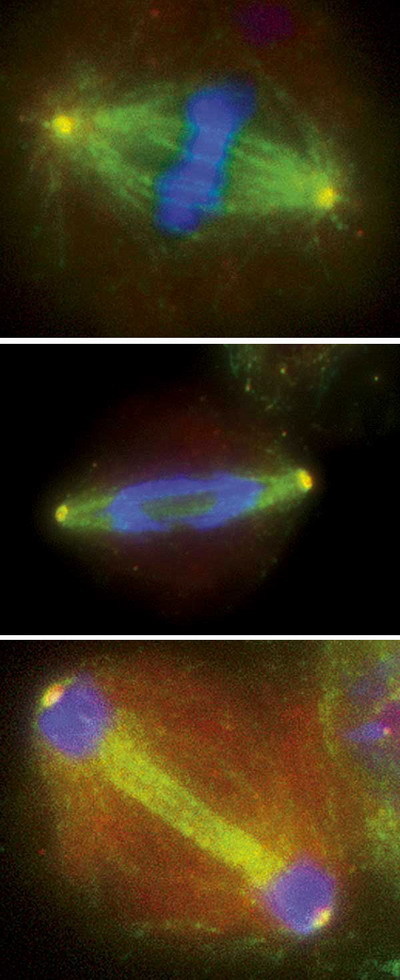
 Co když však jsou oba kinetochory připojeny k mikrotubulům vycházejícím ze stejného pólu? V takovém případě by byly oba chromozomy odtaženy do jedné dceřiné buňky, zatímco druhá by nedostala nic. Podle výsledků získaných v posledních několika letech – zejména díky spolupráci vědců s pivní kvasinkou – se zdá, že buňka umí sledovat napětí mezi mikrotubulem a kinetochorem. Pokud jsou dvě kopie připojeny k mikrotubulům vycházejícím z opačných pólů a zároveň jsou pevně svázány kohezí, působí na ně opačné síly – mikrotubuly se je snaží táhnout od sebe směrem k pólům, zatímco koheze je drží u sebe. V buňce je přítomen protein aurora, který zřejmě dokáže takto vzniklé napětí sledovat, a pokud napětí chybí, odpojí chromozomy od mikrotubulů. Tím dává buňce možnost, aby se o napojení pokusila znova. Tentokrát se jí to snad už povede. Bipolární připojení dvojic chromozomů je také odpovědné za charakteristické uspořádání chromozomů během jedné z fází mitózy, metafázi (obrázek c). Chromozomy se při ní uspořádají krásně přesně uprostřed mezi dvěma póly. Tato pozice je zřejmě fyzikálně nejvýhodnější, neboť tažné síly mikrotubulů vycházející z opačných pólů se vyrovnávají.
Co když však jsou oba kinetochory připojeny k mikrotubulům vycházejícím ze stejného pólu? V takovém případě by byly oba chromozomy odtaženy do jedné dceřiné buňky, zatímco druhá by nedostala nic. Podle výsledků získaných v posledních několika letech – zejména díky spolupráci vědců s pivní kvasinkou – se zdá, že buňka umí sledovat napětí mezi mikrotubulem a kinetochorem. Pokud jsou dvě kopie připojeny k mikrotubulům vycházejícím z opačných pólů a zároveň jsou pevně svázány kohezí, působí na ně opačné síly – mikrotubuly se je snaží táhnout od sebe směrem k pólům, zatímco koheze je drží u sebe. V buňce je přítomen protein aurora, který zřejmě dokáže takto vzniklé napětí sledovat, a pokud napětí chybí, odpojí chromozomy od mikrotubulů. Tím dává buňce možnost, aby se o napojení pokusila znova. Tentokrát se jí to snad už povede. Bipolární připojení dvojic chromozomů je také odpovědné za charakteristické uspořádání chromozomů během jedné z fází mitózy, metafázi (obrázek c). Chromozomy se při ní uspořádají krásně přesně uprostřed mezi dvěma póly. Tato pozice je zřejmě fyzikálně nejvýhodnější, neboť tažné síly mikrotubulů vycházející z opačných pólů se vyrovnávají.
Tak už se konečně rozdělte!
 Ano, je mi jasné, že už jste netrpěliví. Teď už je však dělení opravdu na spadnutí – pokud jsou všechny kopie chromozomů z každé dvojice napojeny na opačně orientované mikrotubuly (v anglickém vědeckém žargonu se tomu říká krásně biorientace), nic už dělení nebrání. No, vlastně brání – koheze! Proteinová pouta držící replikované kopie chromozomů pevně při sobě musí být těsně před poslední fází rozťata. To má na starosti bílkovina separáza. Během celého buněčného cyklu je držena pod kontrolou jinou bílkovinou, zvanou sekurin. Ta zabezpečuje, že se separáza nepustí do kohezinů dřív, než je všechno připraveno. Jakmile jsou všechny chromozomy správně připojeny, buňka dostane od kontrolních proteinů povel, aby se hnula k dalšímu bodu cyklu. Sekurin propustí separázu, která rozstřihne jednu z podjednotek kohezinu, tím koheze povolí a chromozomy jsou na svobodě (obrázek d). Protože jsou připojeny k mikrotubulům, budou ihned po uvolnění přitaženy k pólům, a to vždy každá kopie k pólu jinému (obrázek e). Pak už se vytvoří buněčná přehrádka – a dvě dceřiné buňky, každá s kompletní sadou chromozomů, jsou na světě (obrázek g).
Ano, je mi jasné, že už jste netrpěliví. Teď už je však dělení opravdu na spadnutí – pokud jsou všechny kopie chromozomů z každé dvojice napojeny na opačně orientované mikrotubuly (v anglickém vědeckém žargonu se tomu říká krásně biorientace), nic už dělení nebrání. No, vlastně brání – koheze! Proteinová pouta držící replikované kopie chromozomů pevně při sobě musí být těsně před poslední fází rozťata. To má na starosti bílkovina separáza. Během celého buněčného cyklu je držena pod kontrolou jinou bílkovinou, zvanou sekurin. Ta zabezpečuje, že se separáza nepustí do kohezinů dřív, než je všechno připraveno. Jakmile jsou všechny chromozomy správně připojeny, buňka dostane od kontrolních proteinů povel, aby se hnula k dalšímu bodu cyklu. Sekurin propustí separázu, která rozstřihne jednu z podjednotek kohezinu, tím koheze povolí a chromozomy jsou na svobodě (obrázek d). Protože jsou připojeny k mikrotubulům, budou ihned po uvolnění přitaženy k pólům, a to vždy každá kopie k pólu jinému (obrázek e). Pak už se vytvoří buněčná přehrádka – a dvě dceřiné buňky, každá s kompletní sadou chromozomů, jsou na světě (obrázek g).
Co se může pokazit, to se pokazí
Mitóza je složitý proces, vyžadující dobře načasovanou souhru řady proteinů. Pokud cokoliv selže, dostanou dceřiné buňky buď více, nebo méně chromozomů, než potřebují – budou aneuploidní. Někdy selže dělení buňky kompletně, a místo dvou dceřiných buněk vznikne jedna buňka s dvojnásobnou sadou chromozomů – tetraploidní (obecně polyploidní). Aneuploidie i polyploidie jsou v přírodě překvapivě časté. Podle velkého množství rostlinných druhů, které jsou polyploidní, zřejmě hraje polyploidizace nezanedbatelnou roli v evoluci.Vzhledem k tomu, že se buňka se zvyšující ploidií zvětšuje, je polyploidizace bohatě využívána v šlechtitelství – ty velké jahody ze supermarketu jsou nejčastěji triploidní, nadevropské banány většinou oktaploidní (s osmi sadami chromozomů) a dáte-li si ve Francii u moře krasné velké mušle, budete si pochutnávat na nové sortě (nedávno se slávou uvedené na trh) triploidních a tetraploidních jedinců.
 U živočichů je zvýšení ploidie u celého jedince spíše neobvyklé, přesto se však s následky pokažené mitózy běžně setkáme. Velmi často tvoří aneuploidní či polyploidní buňky rakovinné nádory. Zda je to následek karcinogeneze, či jedna z jejích příčin, je stále ještě předmětem diskusí. Rozhodně je nestabilita chromozomů jedním z charakteristických znaků rakovinných buněk.
U živočichů je zvýšení ploidie u celého jedince spíše neobvyklé, přesto se však s následky pokažené mitózy běžně setkáme. Velmi často tvoří aneuploidní či polyploidní buňky rakovinné nádory. Zda je to následek karcinogeneze, či jedna z jejích příčin, je stále ještě předmětem diskusí. Rozhodně je nestabilita chromozomů jedním z charakteristických znaků rakovinných buněk.
 Buněčné dělení sledované živě pod mikroskopem je úchvatný balet chromozomů, mikrotubulů a dalších buněčných struktur (obrázek, obrázek). Přestože bylo poprvé pozorováno již v předminulém století, stále ještě zůstává mnohé z tohoto procesu utajeno. V posledních letech se však hromadí řada nových poznatků a hlavním motorem tohoto průlomu je mimořádně zdokonalená zobrazovací technika. Mikroskopování se dostalo na novou úroveň, kdy můžeme pozorovat chromozomy, mikrotubuly i další molekuly přímo v akci. A přiznejme si – není lepšího důkazu než vidět něco na vlastní oči.
Buněčné dělení sledované živě pod mikroskopem je úchvatný balet chromozomů, mikrotubulů a dalších buněčných struktur (obrázek, obrázek). Přestože bylo poprvé pozorováno již v předminulém století, stále ještě zůstává mnohé z tohoto procesu utajeno. V posledních letech se však hromadí řada nových poznatků a hlavním motorem tohoto průlomu je mimořádně zdokonalená zobrazovací technika. Mikroskopování se dostalo na novou úroveň, kdy můžeme pozorovat chromozomy, mikrotubuly i další molekuly přímo v akci. A přiznejme si – není lepšího důkazu než vidět něco na vlastní oči.
Kontrola jakosti během buněčného cyklu – CHECKPOINTS
Buňka má řadu kontrolních bodů, kdy ověřuje, jestli všechny buněčné procesy probíhají tak, jak mají. Na základě výsledku jakostní kontroly, která je zajišťována kontrolními bílkovinami (checkpoint proteins), mohou nastat čtyři různé situace:- vše pokračuje podle plánu;
- buňka proces zastaví a čeká, dokud se situace nezlepší;
- buňka proces dokončí, poté vše zastaví a pokusí se napravit, co se pokazilo;
- buňka zkontroluje příslušný proces, zjistí, že je všechno špatně, rozhodne, že by opravy byly příliš náročné či riskantní, a vzdá to – život buňky končí kontrolovanou buněčnou smrtí (apoptózou).
Vědci popisují život buňky od rozdělení k rozdělení jako buněčný cyklus. Během takovéhoto izolovaného buněčného „života“ buňka prochází několika velmi odlišnými fázemi:
- V první fázi, G1, nejdříve buňka zkontroluje, jestli předchozí dělení proběhlo v pořádku a zda obdržela všechny struktury nezbytné k dalšímu cyklu. Během G1 buňka také roste, aby dosáhla velikosti dostatečné k dalšímu rozdělení. Jakostní kontrola v tomto případě zajišťuje, že buňka nezačne s další fází, dokud nedosáhne kritické velikosti.
- Během následující S-fáze zdvojí buňka svou dědičnou informaci procesem replikace. Kontroluje také jak průběh S-fáze, tak její výsledek. Pokud průběžně kvalita replikace klesá, kontrolní bílkoviny replikaci zastaví a nařídí přestávku až do doby, kdy se buňka dá do pořádku tak, aby mohla v replikaci smysluplně pokračovat. V S-fázi se také zdvojují další kritické buněčné struktury, například centrozomy.
- V další fázi, G2, buňka kontroluje, jestli předchozí S-fáze proběhla bezchybně. I v tomto případě zajistí kontrolní bílkoviny přestávku na opravu, pokud je to nezbytně nutné. Během G2-fáze většinou buňka ještě trochu poporoste.
- Následující fáze dělení buňky (M-fáze) je nesmírně komplikovaná a vyžaduje spolupráci řady proteinů. Během této fáze jsou kopie chromozomů rozděleny do dvou sad, obsahujících vždy po jedné kopii od každého chromozomu, a jsou přiděleny vznikajícím dceřiným buňkám. Kontrolní proteiny prověřují, jak probíhá dělení genetické informace, a při každém sebemenším podezření na chybu okamžitě zasahují. Díky tomu dostane každá dcera přesný počet správných chromozomů, které jsou umístěny na opačné póly buňky a odděleny buněčnou přehrádkou. Kontrolní proteiny cyklus zastaví, pokud segregace (oddělování) chromozomů neprobíhá řádně, například jedna buňka by mohla získat o chromozom více, nebo pokud rozmístění dědičné informace neodpovídá plánu, například jsou obě dědičné sady umístěny na stejné straně buňky.

Kromě dohledu na buněčný cyklus zprostředkují kontrolní proteiny také opravy poškození způsobených vnějšími vlivy. Mají toho tedy na starosti mnoho. Musí:
- rozeznat každou chybu,
- zastavit probíhající procesy,
- zastavit i procesy následující,
- rozeznat, kdy byla chyba opravena, a příslušný buněčný proces zase bezchybně nastartovat.
/O mitóze viz též Vesmír 81, 443, 2002/8; Vesmír 75, 505, 1996/9; o chromozomech a genomu Vesmír 70, 323, 1991/6; Vesmír 73, 305, 1994/6; Vesmír 77, 388, 1998/7./
Ke stažení
 článek ve formátu pdf [319,62 kB]
článek ve formátu pdf [319,62 kB]
O autorovi
Zuzana Storchová
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
