










Dobře už bylo
„Biologická rozmanitost Země od prvohor po dnešek roste, ale tento nárůst byl přerušen pěti masovými vymíráními, z nichž to zatím poslední proběhlo před 66 miliony lety a vymřeli při něm například dinosauři.“ S tímto souvětím by asi souhlasila velká část čtenářů Vesmíru, přesto nic z toho neplatí. V posledních letech se zásadně změnily naše představy o dynamice biologické rozmanitosti a tyto poznatky vrhají světlo i na možnou budoucnost biodiverzity v antropocénu.
Převažující náhled na změny biologické rozmanitosti v geologické historii Země byl po několik dekád založen na známé Sepkoskiho křivce, ukazující změny počtu rodů mořských živočichů během fanerozoika (období posledních 540 milionů let, z něhož máme k dispozici slušný fosilní materiál). Ta ukazovala nárůst diverzity na začátku prvohor, potom relativní stabilitu, přerušovanou jen občasnými vymíráními, velké vymírání na přelomu prvohor a druhohor a od té doby trvalý nárůst rozmanitosti, opět přerušený vymíráními (obr. 1A).
Tento obrázek je v posledních dvou dekádách problematizován. Ve Vesmíru jsme už psali o cyklických vzestupech a poklesech diverzity s periodou 62 milionů let, pro kterou se zatím nenašlo uspokojivé vysvětlení (Vesmír 84, 443, 2005/8), i o alternativní křivce vývoje globální diverzity, založené nikoli na interpolacích prvních a posledních nálezů daného rodu, ale na reálném počtu nalezených fosilií z daného období (Vesmír 87, 700, 2008/10). Rigorózní analýza fosilního záznamu, umožněná existencí detailních databází, zproblematizovala ale i věci, na něž dříve paleontologové přísahali, jako je nárůst rozmanitosti směrem k současnosti, a dokonce i populárních pět masových vymírání.
Přerušované rovnováhy biodiverzity
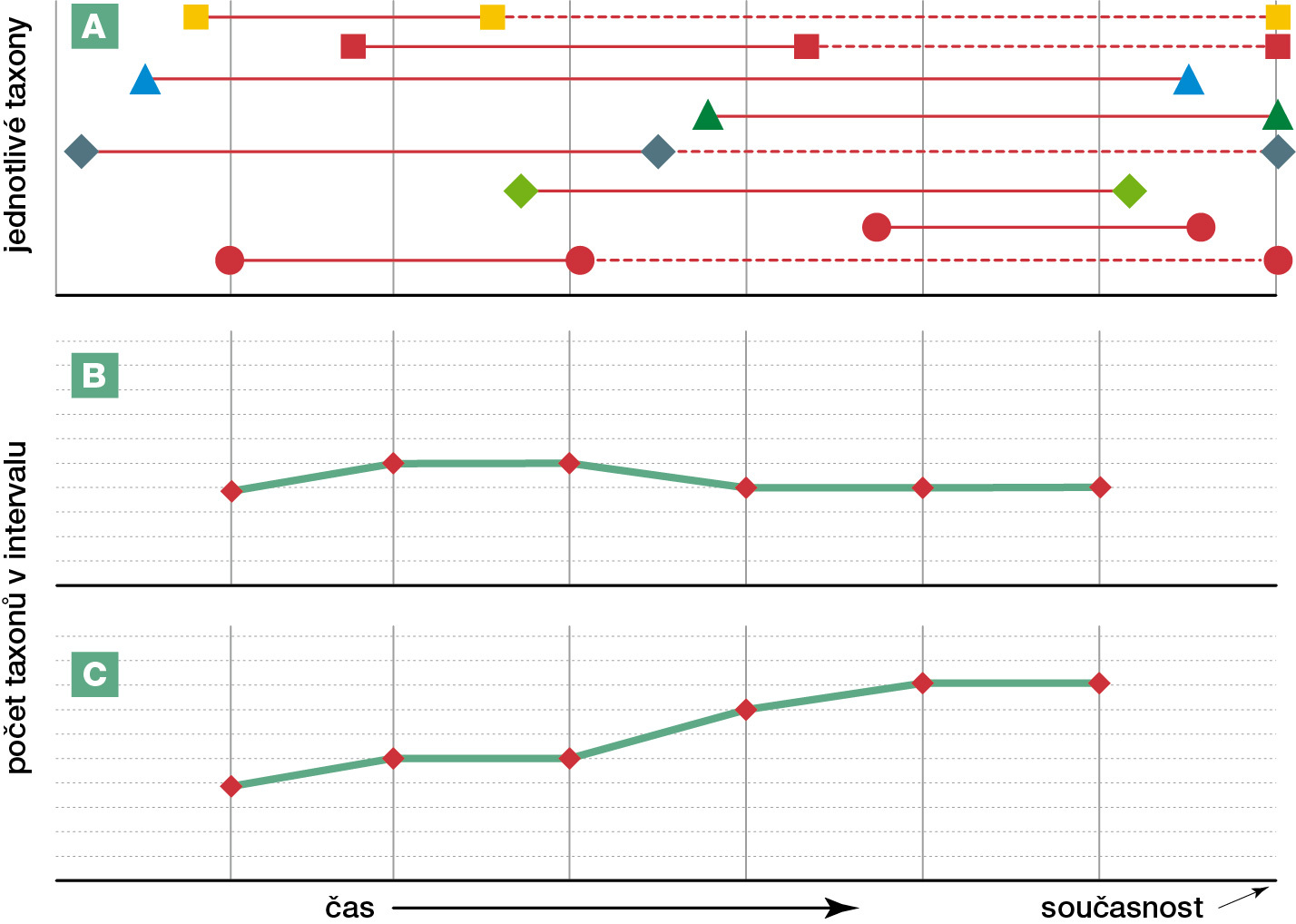
Přestože původní analýzy ukazovaly postupně rostoucí počet taxonů od začátku druhohor až dodnes, už dlouho existovaly pochyby, zda tento nárůst není způsoben tzv. tahem přítomnosti (pull of the recent, obr. 3). Když se tento artefakt eliminoval tím, že se počítaly jen skutečně nalezené fosilie v daném období, a ještě navíc se standardizoval počet vzorků z daných vrstev, vznikla křivka bez zjevného vzestupného trendu, ale dost divoce kolísající (obr. 1B). Nedávno se ale ukázalo, že toto divoké kolísání je do značné míry způsobeno tím, že z různých geologických období máme k dispozici hodně rozdílnou část zemského povrchu, kde vůbec něco fosilizovalo [1]. Problém je, že pokusy tento efekt odfiltrovat mohou zároveň zakrýt reálné kolísání biologické rozmanitosti, poněvadž dostupnost hornin se zkamenělinami může přímo souviset s diverzitou – množství fosilií, které máme k dispozici z moří, hodně souvisí s plochou šelfových moří v tom kterém období, jenže je důvodné předpokládat, že větší rozloha šelfových moří skutečně zvyšovala rozmanitost mořského života. O kolísání globální biologické rozmanitosti v geologické historii toho tedy nelze říci o mnoho více, než že kolísá, snad i trochu periodicky, ale bez zjevného vzestupného trendu.
{kind=link}

S efektem kolísající rozlohy zemského povrchu vhodného k fosilizaci se ale lze vypořádat tak, že se místo na globální diverzitu díváme – zase po řádném statistickém očištění dat – na rozmanitost regionální či lokální. A ta se po dlouhá geologická období mění překvapivě málo. Nejlepší model přitom odpovídá něčemu, co bychom mohli nazvat přerušované rovnováhy, kdyby tento termín nebyl v paleontologii už používán pro něco jiného, totiž pro dlouhodobou neměnnost jednotlivých druhů. Biologická rozmanitost daného území je typicky víceméně stabilní, ale tento rovnovážný stav se mění s velkými přeryvy na rozhraní geologických období [2] – zdá se například, že regionální počet druhů suchozemských obratlovců v druhohorách kolísal kolem nižší hodnoty, než jaká se pak ustavila v kenozoiku (nejmladší období, zahrnující třetihory a čtvrtohory). Biologická rozmanitost určitého území je tedy omezená, ale mění se se zásadními přestavbami celých společenstev a se vznikem evolučních novinek. Jednou takovou novinkou byl zřejmě rozvoj krytosemenných rostlin a na ně navázaného hmyzu během křídy.
Nyní vidíte 19 % článku. Co dál:
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
