










O nepsaných pravidlech, (epigenetické) paměti a (lidském) zapomínání
Molekulární genetika je, jako experimentální obory vědy vůbec, naukou o obecném, reprodukovatelném a spolehlivém. Veškerá osobitost, proměnlivost, rozmanitost a libovůle, k níž mají živí tvorové přirozený sklon prostě proto, že jsou živí, je vnímána jako faktor v lepším případě zanedbatelný, v horším přímo rušivý. Výzkum se soustřeďuje především na pečlivě vybrané modelové organizmy, vyšlechtěné k tomu, aby se chovaly za stejných podmínek všechny stejně, standardně a reprodukovatelně. Opatrnost pak diktuje jedno z nepsaných základních pravidel experimentální genetické práce: chová-li se část zkoumaného systému v každém pokusu jinak, je lépe jej zahodit než se s ním zahazovat. Ne že by takový systém nemohl být zcela legitimním předmětem zkoumání – prostě s sebou nese zvýšené riziko, že se nedostaví výsledky a nebudou publikace, což může být pro vědcovu kariéru osudné.
S nestabilním systémem ale snadno zahodíme i šanci objevit ve zdánlivé nepravidelnosti pořádek, který stojí za další zkoumání a který může vypovídat o něčem důležitém, co by se jinak dalo sotva tušit. Nedbat nepsaných pravidel je však nebezpečné. Badatel, který rozhoduje mezi „zahozením“ a „zahazováním se“, volí zároveň mezi jistotou zavedené univerzitní existence a dobrodružstvím setkávání s neznámem.
Útěchu snad lze nalézt ve skutečnosti, že rozhodování, ve kterém jsou v sázce jen budoucí publikace, je pouhou malicherností ve srovnání se situacemi, v nichž jde doslova o život. Ne, nehodlám zde moralizovat a připomínat drastické příběhy z dějin či současnosti lidstva. Životně důležitá rozhodnutí totiž nečiníme jen my: není snad každé zapnutí či vypnutí genu, na jehož činnosti závisí budoucí osud buňky, také svého druhu volbou, která může mít dalekosáhlé důsledky?
To, jak důležitá rozhodnutí se mohou odehrávat na úrovni řízení aktivity genů, lze ilustrovat klasickým učebnicovým příkladem, jehož „hrdinkou“ je ochočená střevní bakterie Escherichia coli a virus, který na ní parazituje. Dnes se s E. coli můžeme setkat na pracovním stole kteréhokoli molekulárního biologa, jenž se zabývá genovým inženýrstvím a buňky této bakterie používá jako nenáročné a poměrně spolehlivé „živé zkumavky“ k uchovávání a pomnožování kousků DNA či k přípravě proteinů. Pověstná spolehlivost E. coli je ale výsledkem dlouholeté usilovné selekce kmenů a optimalizace podmínek. Za jiných okolností a v jiném experimentálním uspořádání se naše bakterie chová jako plnohodnotný živý tvor se vším všudy – včetně toho, že může „onemocnět“, tedy být parazitována viry, které se označují jako bakteriofágy či prostě fágy.
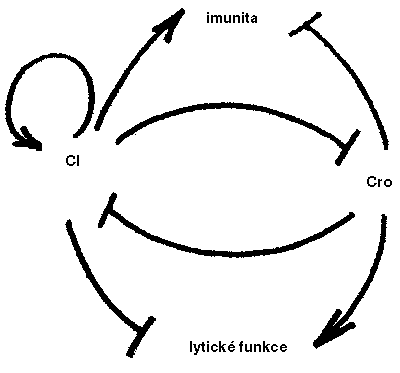
Napadení bakteriální kultury fágem se v nejjednodušším případě projevuje tak, že infikované bakterie zahynou a rozpadnou se za současného uvolnění velkého počtu nových fágových částic, které infikují další bakterie (obrázek). Protože postižené bakterie hynou a rozpadají se (lyzují), hovoříme o lytické infekci fágem. Některé fágy, například bakteriofág lambda (λ), však hostitelské bakterie zabíjejí jen někdy. Kromě lytické infekce mohou totiž vyvolat i infekci lyzogenní – místo toho, aby se fág pomnožil a zároveň zničil hostitele, zabuduje svůj genom do hostitelského a tiše přetrvává v latentním stavu jako profág. Může se přitom „probudit“ zpět do lytického stavu. To se stává zejména v případě, že hostitelská bakterie byla nějak poškozena, třeba ozářením ultrafialovým světlem (lze snadno nahlédnout, že probuzení profága v poškozené a možná umírající bakterii je záležitostí veskrze adaptivní). Z hlediska fága je lyzogenní stav docela dobrým prostředkem jak si zajistit přetrvání a pomnožení téměř bez námahy s vyráběním fágových proteinů. Z celého profága λ je aktivní pouze jediný gen, jehož proteinový produkt – represor CI – aktivuje svůj vlastní gen a současně udržuje všechny ostatní fágové geny ve „vypnutém“ stavu. I pro bakterii je výhodné vydržovat si profága – poměrně malá zátěž plynoucí z produkce jednoho fágového proteinu je více než kompenzována tím, že lyzogenní bakterie je imunní vůči lytické infekci stejným fágem.
{kind=link}
Fág, jenž právě infikoval „čistou“ buňku E. coli, se tedy rozhoduje mezi dvěma životními strategiemi, které se vzájemně vylučují: buď zapne gen pro represor CI a stane se profágem v lyzogenní buňce, anebo zapne gen pro antirepresor kódovaný genem ai/cro, který zabrání aktivaci genu CI a umožní spuštění kaskády genů potřebných pro úspěšný průběh lytického životního cyklu, zabití hostitele a pomnožení fága (obrázek). 1) Klíčovou roli v tomto rozhodování hraje specifická sekvence DNA, která je schopna vázat jak represor, tak antirepresor. Podle toho, který z proteinů obsadí toto regulační místo, zapne se buď CI (bakterie se stane lyzogenní), nebo ai/cro (fág se pomnoží a bakterie zahyne). Do poměru koncentrací CI a ai/cro se promítá řada faktorů – například fyziologické nastavení hostitelských buněk a jejich zdravotní stav či počet fágových částic, které bakterii infikovaly. Tento „genetický přepínač“ s dvěma regulačními stavy se stal přímo prototypem buněčného rozhodovacího mechanizmu schopného integrovat a vyhodnocovat řadu různorodých vstupů; v nějaké podobě na něj narazíme téměř v každé učebnici molekulární genetiky.
{kind=link}
Méně často však už se učebnice zabývají tím, jak se na celou záležitost přišlo. Je to škoda – nejen pro ty z nás, kteří mají v lásce a úctě Zdeňka Neubauera, jehož práce z přelomu 60. a 70. let byly zcela zásadní pro identifikaci antirepresoru a pochopení funkce „genetického přepínače“ (viz zejména Neubauer, Nature 213, 1263, 1967; Calef a Neubauer, Cold Spring Harbor Symp. Quant. Biol. 23, 765, 1968; Oppenheim, Neubauer a Calef, Nature 226, 31, 1970; Neubauer a Calef, J. Mol. Biol. 51, 1, 1970, Hampacherová, Koutecká a Neubauer, Genetics 120, 133, 1973). V učebnicích totiž bývají klasické experimenty popisovány většinou jen tehdy, když jsou jednoduché, dají se shrnout na půl stránky a jeden obrázek ...a neuvádějí v pochybnost nepsaná pravidla řemesla. Ani jedno z toho naneštěstí o dotyčných pokusech neplatí, a proto se čtenáři učebnic musí spokojit se suchým shrnutím výsledků. Zůstanou sice ušetřeni metodických detailů a komplikované terminologie fágových mutantů, ale také úžasu, který se zmocňuje autorky těchto řádků při zjištění, jak to vlastně bylo.
Na začátku byla příprava speciálního kmene lyzogenních bakterií s poškozeným represorem CI, který ve zvýšené teplotě přestává fungovat. Profág by tedy měl přejít do lytické fáze životního cyklu a zahubit hostitelské buňky za současného pomnožení fága. Dodatečnou mutací fágového genu nutného pro první kroky lytické fáze však lze množení fága a lyzi buněk zabránit. Bakterie pak přežívají pobyt ve vysoké teplotě, který by indukoval lytické množení fága – a můžeme na nich studovat otázku, která by za normálních okolností neměla smysl: jsou buňky po neúspěšné indukci lytické fáze imunní vůči další infekci? Odpověď se ukázala být přímo šokující: záleží na okolnostech. Po několika generacích bez funkčního represoru buňky ztrácejí imunitu, a dokonce získávají jistou predispozici k lytickému průběhu infekce – a neimunní stav se může dlouhodobě (dědičně) udržovat. Dnes víme, že je to proto, že si bakterie stačily vytvořit antirepresor, a ten obsadil vazebná místa pro represor. V populaci neimunních bakterií se však záhy spontánně objevuje několikaprocentní frakce buněk s obnovenou imunitou. Nejsou to mutanti: při dlouhodobém udržování v kultuře produkují tyto imunní klony naopak malou frakci buněk neimunních, a podobně jako výchozí kulturu je můžeme zbavit imunity vystavením zvýšené teplotě. (Při současných znalostech je pozoruhodná nestabilita defektních lyzogenů pochopitelná – stačí si uvědomit, že o osudu buňky rozhoduje doslova několik molekul, a tudíž se občas stává, že některá buňka při dělení podědí příliš málo represoru či antirepresoru). Celý postup lze po libosti opakovat, bakteriální kultura jako by procházela jakýmsi fázovým přeskokem mezi dvěma regulačními stavy.
Právě objev dvou stabilních regulačních stavů jistých mutantů fága λ byl jedním z prvních kroků k popisu „genového přepínače“ CI – ai/cro. Zároveň však jde o zjištění podstatně obecnějšího a velice zásadního dopadu: totiž o to, že dědičná paměť organizmu může být „zapsána“ i epigeneticky – to jest jinde než v posloupnosti nukleotidů DNA. Regulační stavy genů se mohou dědit přes mnoho generací stejně jako geny samy. Jde však o zásadně, kvalitativně jiný druh paměti, plastičtější, než je „tvrdá“ paměť zapsaná v sekvencích DNA. Proto se to také tak těžko studuje: není snad lepší příklad systému, který je radno zahodit, abychom se s ním nezahazovali, než populace bakterií, která neustále odštěpuje buňky odlišných vlastností!
Nechce se mi věřit, že by na přelomu let šedesátých a sedmdesátých, kdy Zdeněk Neubauer v Calefově laboratoři v Neapoli, a posléze na katedře genetiky a mikrobiologie pražské Přírodovědecké fakulty studoval fágy, býval vědecký svět nějak zásadně jiný než ten náš dnešní. Volba mezi „zahozením“ a „zahazováním se“ byla zajisté už tehdy rozhodováním mezi snad trochu nudnou, ale bezpečnou jistotou na jedné straně – a nádherným, byť i riskantním dobrodružstvím poznání na straně druhé. Jak Zdeňka znám, jistě dlouho neváhal. Koneckonců i jeho pozdější rozhodnutí věnovat se naplno filozofii bylo podobným krokem do temného neznáma, kde ani největší optimista tehdy nemohl tušit budoucí profesuru.
Samozřejmě mohlo být i jinak.
Ale to by vlastně nebylo proč psát... 1)
Poznámky
Ke stažení
 Článek ve formátu PDF [495,62 kB]
Článek ve formátu PDF [495,62 kB]
O autorovi
Fatima Cvrčková

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
