










Stresové proteiny v imunitě
Co nevyléčí léky, vyléčí skalpel.
Co nevyléčí skalpel, vyléčí teplo.
Hippokrates
Stresové proteiny jsou jedny z nejstarších bílkovin vůbec. Vznikly pravděpodobně s prvními buňkami v horkém prostředí, které je dnes domovem teplomilných bakterií. Brzy se naučily vyvarovat se příliš vysokých teplot, které by je mohly zahubit. A když už to uměly, staly se užitečnými strážci ostatních bílkovin, které to nedovedou. O jednotném původu života svědčí skutečnost, že všechny organizmy mají nejen nukleové kyseliny a proteiny, ale i více než 93 % společných metabolických drah. To ale ještě není všechno. Společné jsou totiž i molekulární mechanizmy, jimiž se buňky brání proti různým formám stresu.
Odpověď vyvolaná stresem zahrnuje aktivaci určitých genů pro takzvané stresové proteiny čili bílkoviny teplotního šoku, známé pod anglickým názvem heat shock proteins (HSP). Termín bílkoviny teplotního šoku se původně užíval pro všechny proteiny, jejichž tvorba vzrostla bezprostředně po působení tepla. Ukázalo se však, že některé z těchto proteinů jsou v buňce syntetizovány stále, vyšší teplota jejich produkci pouze podstatně zvýší.
Stopa z archaika
Představme si scenérii archaika, nejstaršího období (před třemi až čtyřmi miliardami let). Oranžové slunce na růžové obloze vytvořené atmosférou dusíku se stopami kysličníku uhličitého ozařovalo hnědý oceán. V něm zdánlivě beznadějně zápasily o přežití první živé buňky, jež pomocí reakce uhličitanu železnatého se sirovodíkem získávaly energii pro svůj metabolizmus. Nepotřebovaly tolik energie jako dnešní organizmy. Žily ve výhodném horkém prostředí, které je však mohlo svou vysokou teplotou usmrtit. Potřebovaly se vyhnout místům, kde teplota přesahovala 100 °C, a proto si vytvořily čidlo citlivé na infračervené záření. Detektorem mohly být například bakteriální chlorofyly (z nich se později vyvinul zelený pigment rostlin). Pokud byly buňky krátkodobě vystaveny vyšším teplotám, použily na svou ochranu systém stresových proteinů. S určitou nadsázkou lze říci, že tyto bílkoviny zajistily přežití biosféry a že právě jim vděčíme za svůj život.
Průvodci bílkovin
Úloha bílkovin teplotního šoku v buňce je mnohostranná. Základním úkolem těchto bílkovin však je ochrana správného prostorového uspořádání ostatních buněčných proteinů. Jak to dělají? Zabraňují změnám molekulární struktury u ostatních bílkovin, a tím je chrání před denaturací a ztrátou funkčnosti. Působí vlastně jako strážci a průvodci ostatních bílkovin. (Odtud pocházejí jejich další názvy chaperony a chaperoniny, které vznikly z francouzského un chaperon [šaper] – gardedáma, viz také Vesmír 77, 15, 1998/1). Když na buňky působí nějaká vnější zátěž (teplotní změny nebo osmotický šok, měnící se chemizmus prostředí apod.), provázejí strážci polypeptidy přes různá úskalí při pouti buňkou a střeží neporušenost jejich struktury při přestupu přes membrány nebo při vstupu do buněčných organel (například mitochondrií). Proteiny teplotního šoku však hrají významnou úlohu i za nestresových podmínek ve vývojových stadiích buňky, v buněčném cyklu, a tím ve výstavbě a funkci tkání.Pro objasnění jejich významu si můžeme dovolit určitou analogii. Stejně jako imunitní buňky udržují integritu vnitřního prostředí mnohobuněčného organizmu, plní „proteinoví hlídači“ integrační funkce ve vnitřním prostředí buňky. Tím, že brání narušení buněčných funkcí, udržují i funkčnost tkání a orgánů a mají zásadní význam pro přežití organizmu jako celku.
Buňky v neživém světě
Buňky jsou křehké soustavy. Představme si jejich nepatrnost oproti vesmírným a planetárním měřítkům a na druhé straně drsnost a agresivitu fyzikálně-chemického světa, v němž vznikaly, vyvíjely se, začaly vzájemně komunikovat a spojovat se, až vznikly mnohobuněčné organizmy. Co umožnilo jejich přetrvání? V současné době je možná jediná odpověď: buňky si již v dobách svého vzniku vytvořily speciální molekuly k obraně a útlumu nepříznivých vlivů vnějšího světa (viz obrázek).V šedesátých a sedmdesátých letech provedl C. B. Anfinsen se spolupracovníky laboratorní experimenty, jimiž prokázal, že veškerá informace o struktuře a konečné konfiguraci molekuly polypeptidu je obsažena v samotném polypeptidickém řetězci (Anfinsenovo dogma). To znamená, že se denaturovaný enzym může složit do původního prostorového uspořádání bez pomoci jiných proteinů. Ukázalo se však, že podmínky „ve zkumavce“ neodrážejí přesně to, co se děje v živé buňce, kdy právě syntetizovaná molekula proteinu zaujímá svou konečnou prostorovou uspořádanost. Anfinsenovo dogma in vivo neplatí. V prostředí buňky, která je doslova přeplněna různými a různě reagujícími druhy molekul, vyžaduje skládání proteinového řetězce přítomnost pomocných molekul – chaperonů.
Řízení buněčné stresové odpovědi
Prozatím nejvíce víme o tom, jak probíhá odpověď na stres z přepisu genů. Vysoce konzervativní oblast DNA zvaná element teplotního šoku (heat shock element, HSE) se naváže na specifický transkripční faktor (HSF). Když se element teplotního šoku odstraní, vede to k neschopnosti buňky odpovědět na stres, a když se naopak geneticko-inženýrskou manipulací zavede zpátky, buňce se zmíněná schopnost zase vrátí.Specifický transkripční faktor aktivuje přepis genů kódujících bílkoviny teplotního šoku. U kvasinek a octomilky je prozatím znám jeden transkripční faktor, u rajčete a kura tři, u savců dva.
V roce 1992 Richard I. Morimoto se spolupracovníky navrhl model autoregulace stresové odpovědi, který vysvětluje, jak transkripční faktor účinkuje. V klidové buňce je vázán na HSP70 (protein teplotního šoku ze skupiny stresových proteinů o relativní molekulové hmotnosti kolem 70 000). Teplotní nebo jiné formy stresu zvýší počet poškozených nebo denaturovaných proteinových molekul, které s transkripčním faktorem soutěží o vazbu na HSP70. Transkripční faktor se uvolní a vstoupí do jádra, kde aktivuje přepis genů proteinu teplotního šoku. Jakmile je dosaženo hladiny transkripčního faktoru dostatečné k tomu, aby se volné poškozené proteiny vyvázaly, opět se koncentrace transkripčního faktoru zpětnou vazbou sníží a stresové reakce ustupují.
Stresové proteiny slouží hlavně k tomu, že odklízejí poškozené a chybně složené bílkovinné molekuly v bakteriálních, rostlinných i živočišných buňkách, a v živočišných buňkách také stabilizují a přetvářejí vnitřní buněčnou kostru (cytoskelet).
Rodiny stresových proteinů
Bílkoviny teplotního šoku se obvykle dělí podle molekulární hmotnosti do několika rodin. Počet nově identifikovaných proteinů každý měsíc vzrůstá. Zatím nejmenší bílkoviny teplotního šoku mají relativní molekulovou hmotnost 27 000 a uplatňují se při přenosu signálů cytokinů, molekul, které regulují činnost imunitních buněk. Známou molekulou je ubikvitin, který provází denaturované proteiny do speciálních pohřebišť – proteazomů (viz Vesmír 75, 250, 1996/5). Větší bílkoviny teplotního šoku (o relativní molekulové hmotnosti 47 000) se účastní tvorby a sekrece kolagenu (mezibuněčné hmoty, z níž se vyrábí klih; viz Vesmír 75, 429, 1996/8). Největší molekuly stresových proteinů mají relativní molekulovou hmotnost 110 000. Protein vázající imunoglobuliny má hmotnost 78 000 a jeho produkce se zvyšuje při nedostatku glukózy. Váže těžký řetězec imunoglobulinové molekuly a přenáší právě vytvořené polypeptidy do endoplazmatického retikula. Nejzajímavější stresové proteiny mají relativní molekulovou hmotnost 65 000–70 000. Podívejme se na ně blíže.Rozpoznání antigenu
Při pohlcování odumřelých červených krvinek se v lidských fagocytech zvyšuje syntéza proteinů, které působí jako doprovod (chaperony) a přispívají k zneškodnění produktů fagocytárního rozpadu.
Byla prokázána spojitost genů stresových proteinů na šestém lidském chromozomu s hlavním histokompatibilitním komplexem (ř. histos – tkáň, l. compatí– spolucítit), který určuje tkáňovou snášenlivost či neslučitelnost v transplantační imunitě.
V r. 1985 vyslovil D. R. Forsdyke hypotézu o neimunologickém rozpoznávání vlastních a cizích antigenů, v němž právě tyto proteiny mohou hrát významnou úlohu při obraně proti nitrobuněčným parazitům. Její podstatou je rozpoznávání nesprávně strukturovaných nebo abnormálních proteinů. Takto „označené“ antigeny jsou pohlceny makrofágem či dendritickou buňkou, které podivný protein naštípou na krátké oligopeptidy schopné vazby na molekulu hlavního histokompatibilitního komplexu. Ta předloží antigenní oligopeptid lymfocytu T a tento jev (prezentace antigenu) je počátkem specifické imunitní odpovědi. Lymfocyty T totiž nejsou schopny rozpoznat antigen v jiné formě než ve vazbě na membránovou molekulu histokompatibilního komplexu na buňce, kde je antigen prezentován.
Stresové proteiny v infekci
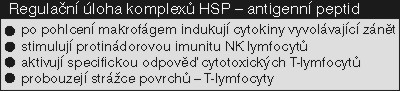
HSP70 aktivují produkci cytokinů – signálních molekul imunity (interleukinu-12). Samy jsou aktivovány enzymy zvanými proteinkinázy SAPK, jež jsou zase indukovány cytokiny. Aby imunitní odezva měla zpětnovazební regulaci, HSP70 brání sekreci důležitého prozánětového cytokinu TNF (faktoru nekrotizujícího nádory). Přítomnost stresového proteinu 70 je charakteristickým průvodním znakem bakteriálních, virových a mykotických infekcí a je typická u zánětu – u akutního krátkodobá, u chronických zánětů trvalá (viz také obrázek).
Stresový protein 70 a revma
Strukturální podobnost molekul lidských a bakteriálních stresových proteinů 70 dosahuje asi 50 %. To má za následek, že některé sledy aminokyselin v těchto molekulách jsou totožné a že se v těle mohou namnožit klony lymfocytů, které je rozpoznávají a začnou reagovat na vlastní bílkoviny jako na bílkoviny bakterií. Většinou je utlumena imunitní odpověď, ale v mnoha případech se tato tolerance nevytvoří, rozjede se imunitní reakce proti vlastní tkáni, a posléze vznikne autoimunitní onemocnění. Z hlediska zdraví jde o velmi závažný jev. Předpokládá se, že se tato systémová chyba imunitní soustavy uplatňuje při vzniku revmatického onemocnění kloubů a snad i dalších nemocí. Vzhledem k četnosti revmatu je význam stresových proteinů v medicíně obrovský. Ukázalo se, že rekombinantní stresový protein 70 zabránil vzniku pokusného revmatického onemocnění u myší. Potvrdil se tak smysl studia těchto molekul, z nichž promlouvá sama historie počátku života na naší planetě. 1)Poznámky
Citát
Stanisłav Lem v knize Pavla Weigla: Stanisłav Lem, Magnet-Press, Praha 1995, s. 87-88
…domnívám se, že tak „normální“ literatura, jakou je science fiction, je každým rokem stále ohroženější. Toto ohrožení nevyplývá z toho, že vznikla „ohromná konkurence“ v podobě filmu, televize, masových sdělovacích prostředků, že lidé raději hledí na velkou či malou obrazovku, než aby četli knihy. Toto ohrožení pochází z toho, že nedokážeme rozeznat jejich reálnou hierarchii – rozlišit odstupňování jejich závažnosti pro náš okamžitý i budoucí osud. Pociťujeme, že civilizace v prudkém pohybu se odtrhává, že je odtrhávána od svých tradičních, historických kořenů, že proto musí sondovat svou budoucnost, že musí dnes dělat rozhodnutí, jejichž důsledky zachrání nebo zahubí naše děti a vnuky. Tento stav přesahuje naše síly a někdy se mu říká future shock, tedy ochrnutí z vizemi nepostižitelné, rozpory rozervané a současně nezadržitelně přicházející budoucnosti. Tento stav zastihl literaturu i science fiction nepřipravené. To, o čem dnes vypráví „normální“ beletrie, i to, co vypravují barvité knihy SF, se rozchází se světem, který existuje, a ještě více se světem, který přichází branami vedoucími do 21. století. „Normální“ literatura se často uzavírá do sebe nebo přechází do mytologických pohádek nebo nás straší zjednodušenými vizemi budoucích civilizačních děsů. Obě se tedy uchylují k nesprávným formám – rezignují z jednání, které dalo literatuře důstojnost a které J. Conrad nazval „vyměřování spravedlnosti viditelnému světu“. Aby však bylo možné vyměřit spravedlnost, je nejdřív potřeba pochopit důvody souzených stran; nemůže tedy být nic důležitějšího než úsilí pochopit, kam náš svět směřuje a zda tomu máme klást odpor, nebo tento pohyb akceptovat a aktivně se jej účastnit.
STRESOVÝ PROTEIN 70
- Ve vývoji druhu: HSP70 se vyskytují u všech živých organizmů a představují jednu z nejkonzervativnějších proteinových rodin vůbec. Díky svému rozšíření a vysokému stupni strukturální homologie slouží pro studium vývojové příbuznosti živých organizmů. Morfologická a biochemická podobnost buněčných organel, jako jsou mitochondrie nebo chloroplasty, s určitými druhy eubakterií dala podnět k domněnce 2) o původu organel ze symbiotických bakterií. Domněnku potvrdily právě klonační a sekvenční studie HSP 70.
- Ve vývoji jedince: Časná embrya jsou na teplotní změny citlivá, protože na ně nemohou reagovat stresovou buněčnou odpovědí typickou pro dospělé organizmy. Termosenzitivita embryí byla prokázána u tak odlišných druhů, jako je octomilka, mořská ježovka a myš! Pravděpodobně je způsobena nepřítomností transkripčního faktoru 2 v určitém stadiu vývoje, což by mohlo naznačovat významnou roli stresových proteinů v časné embryogenezi.
HSP70 se objevuje bez vnějšího zásahu v embryonálních buňkách během vzniku gastruly. Toto časné zárodečné stadium se z oplozeného vajíčka vyvíjí asi po čtyřech dnech (kdy už je vajíčko rozrýhováno na množství zárodečných buněk – blastomer). Způsob vzniku je podobný promáčknutí stěny vyfouknutého míče (viz Vesmír 76, 89, 1997/2). HSP70 je v buňkách embrya i při vytváření nervové trubice a oka.
HSP65 – savčí homolog chaperoninu GroEL střevní bakterie Escherichia coli byl prokázán v embryonálních tkáních člověka, prasete a myši. Například v prasečích embryonálních játrech se tato molekula objevuje v období maximální krvetvorby.
Ke stažení
 Článek ve formátu PDF [138,46 kB]
Článek ve formátu PDF [138,46 kB]
O autorech
Petr Šíma
Ilja Trebichavský
Doporučujeme


{kind=link}
{kind=link}
{kind=link}