










Prvoci kmene Myxozoa novou skupinou vícebuněčných živočichů
| 5. 3. 1996Zavedení nové metody v jakémkoli vědeckém odvětví může nejen zpřesnit dosavadní znalosti, ale i úplně zpřeházet existující představy. V biologii jsou skupinou, která je více než jiné závislá na metodickém pokroku, protisti. Zopakujme pro osvěžení paměti, že tvoří říši eukaryotických organizmů, vybavených již pravým buněčným jádrem uvnitř dvojité jaderné membrány, že jsou až na výjimky mikroskopických rozměrů a stojí u začátku vývojových linií vedoucích k houbám, rostlinám a živočichům. Prvoci (Protozoa) pak jsou tradičně vžitý, ale zcela arbitrární termín pro jakousi těžko vymezitelnou „podříši“ protistů spíše živočišného typu. Některé z nich pozoroval sice už A. Leeuwenhoek v 17. století, ale ostatní museli počkat až na skutečný rozvoj světelné mikroskopie. Ta umožnila poznat jejich obrovskou rozmanitost, početnost a skoro všudypřítomnost, uspořádat je do stále vylepšovaných systémů a získat jakousi představu o jejich fylogenezi. Opravdovou revolucí v poznávání jejich struktur a příbuzenských vztahů byl nástup elektronové mikroskopie v padesátých letech.
Toto objevování stále ještě neskončilo a trvá, i když už i do protistologie vtrhla molekulární biologie. Pronikla do všech směrů výzkumu a klíčový význam má i při objasňování fylogeneze a příbuzností protistů mezi sebou a ve vztahu k ostatním organizmům. Využívá se k tomu zejména rozdílů v sekvencích nukleotidů a změn, ke kterým docházelo v průběhu evoluce v genu kódujícím tzv. malou podjednotku ribozomální RNA, označovanou jako 18s rRNA. Tento gen je zastoupen u všech organizmů počínaje bakteriemi a umožňuje nejširší srovnávání evoluční vzdálenosti organizmů a tím i sestrojení jejich pravděpodobného rodokmenu. U protistů tak bylo dosaženo překvapivých zjištění, že nikoli jednoduší volně žijící bičíkovci, ale strukturálně velmi složití cizopasní prvoci rodu Giardia a mikrosporidie jsou mezi protisty nejblíže k bakteriím, a tedy vůbec na nejnižším stupni vývoje všech eukaryotických organizmů. K ještě pozoruhodnějším závěrům ale dochází molekulární biologie u kmene Myxozoa, jednoho velkého kmene protistů.
Myxozoa

 Tento kmen tvoří velkou, výlučně parazitickou skupinu organizmů se skoro 1 300 druhy řazenými do 53 rodů. Člení se do dvou tříd, Myxosporea s druhy, které cizopasí převážně u ryb, a Actinosporea, což je malá a málo známá třída s pouze asi 40 popsanými druhy, které žijí především v kroužkovcích (Annelida). Proto pokud hovoříme o myxozoích, máme většinou na mysli zástupce první třídy, myxosporidie. Ty působí v orgánech ryb často velmi nápadné změny - tvoří i velká ložiska, běžně patrná jako bílé uzlíky plné spor (viz obrázek a obrázek), nebo prostupují drobnými vývojovými stadii celé orgány těla. Mohou působit vážná onemocnění i úhyny ryb, nebo přivodí taková znetvoření, že ryby jsou neprodejné.
Tento kmen tvoří velkou, výlučně parazitickou skupinu organizmů se skoro 1 300 druhy řazenými do 53 rodů. Člení se do dvou tříd, Myxosporea s druhy, které cizopasí převážně u ryb, a Actinosporea, což je malá a málo známá třída s pouze asi 40 popsanými druhy, které žijí především v kroužkovcích (Annelida). Proto pokud hovoříme o myxozoích, máme většinou na mysli zástupce první třídy, myxosporidie. Ty působí v orgánech ryb často velmi nápadné změny - tvoří i velká ložiska, běžně patrná jako bílé uzlíky plné spor (viz obrázek a obrázek), nebo prostupují drobnými vývojovými stadii celé orgány těla. Mohou působit vážná onemocnění i úhyny ryb, nebo přivodí taková znetvoření, že ryby jsou neprodejné.
 Myxosporidie jsou charakterizovány svými infekčními stadii, sporami, které vznikají uvnitř mateřského plazmodia z několika buněk. Některé se přetvoří na odolné skořápky spory, které obalují buňku vlastního infekčního zárodku, sporoplazmy, jiné buňky vytvářejí pólové váčky umístěné na jednom nebo dvou pólech spory (viz obrázek). Tyto silnostěnné, pod mikroskopem světlolomné váčky mají v sobě dlouhé, spirálně stočené vlákno, které je schopno doslova bleskového vymrštění (obrázek), podobně jako je tomu u žahavých buněk (nematocyst) žahavců, například nezmarů či medúz. Podle typu nematocyst má vymrštěné vlákno různé funkce.
Myxosporidie jsou charakterizovány svými infekčními stadii, sporami, které vznikají uvnitř mateřského plazmodia z několika buněk. Některé se přetvoří na odolné skořápky spory, které obalují buňku vlastního infekčního zárodku, sporoplazmy, jiné buňky vytvářejí pólové váčky umístěné na jednom nebo dvou pólech spory (viz obrázek). Tyto silnostěnné, pod mikroskopem světlolomné váčky mají v sobě dlouhé, spirálně stočené vlákno, které je schopno doslova bleskového vymrštění (obrázek), podobně jako je tomu u žahavých buněk (nematocyst) žahavců, například nezmarů či medúz. Podle typu nematocyst má vymrštěné vlákno různé funkce. 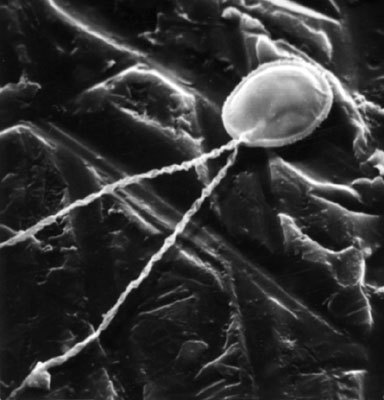 Nejznámější jsou žahavá vlákna, která vylučují toxiny omračující kořist žahavců. Připletou-li se k trubýšům nebo medúzám, jejichž chapadla jsou posázena žahavými nematocystami, lidé koupající se v moři, mohou jimi být velmi těžce popáleni. U myxosporidií však vystřelitelná vlákna pólových váčků mají jediný zřejmý účel - přichycení ke stěně zažívací roury infikovaného hostitele při otvírání spory. Vymrštění vláken je možné také navodit uměle nejrůznějšími chemickými a osmotickými stimuly. Stejně jako spory jsou složitá i vegetativní stadia myxosporeí, tvořící drobná i velká, často makroskopická plazmodia v tkáních i orgánových dutinách ryb. Plazmodia obsahují kromě svých jader i tzv. generativní buňky. Ty „plavou“ v cytoplazmě plazmodia a jejich sdružováním, dělením a diferenciací pak vznikají spory. Ve velkých tkáňových plazmodiích, jaká jsou častá na žábrách ryb, jich mohou vzniknout desítky nebo i stovky tisíc.
Nejznámější jsou žahavá vlákna, která vylučují toxiny omračující kořist žahavců. Připletou-li se k trubýšům nebo medúzám, jejichž chapadla jsou posázena žahavými nematocystami, lidé koupající se v moři, mohou jimi být velmi těžce popáleni. U myxosporidií však vystřelitelná vlákna pólových váčků mají jediný zřejmý účel - přichycení ke stěně zažívací roury infikovaného hostitele při otvírání spory. Vymrštění vláken je možné také navodit uměle nejrůznějšími chemickými a osmotickými stimuly. Stejně jako spory jsou složitá i vegetativní stadia myxosporeí, tvořící drobná i velká, často makroskopická plazmodia v tkáních i orgánových dutinách ryb. Plazmodia obsahují kromě svých jader i tzv. generativní buňky. Ty „plavou“ v cytoplazmě plazmodia a jejich sdružováním, dělením a diferenciací pak vznikají spory. Ve velkých tkáňových plazmodiích, jaká jsou častá na žábrách ryb, jich mohou vzniknout desítky nebo i stovky tisíc.
Prvoci, nebo mnohobuněční?
Druhy napadající ryby byly objeveny poprvé už r. 1825 u síhů v Ženevském jezeře ve Švýcarsku, tedy mnohem dříve než nejznámější cizopasní prvoci, jako třeba původci malárie nebo spavé nemoci. U myxosporidií, poněvadž napadají „pouze“ ryby, postupovala prvá (světelněmikroskopická) etapa poznávání mnohem pomaleji. Teprve v prvých desetiletích tohoto století byla získána představa o jejich biologii. Prokázalo se, že se přenos děje z ryby na rybu prostřednictvím spor uvolněných do vody, které nový hostitel pozře. V jeho střevě se spora otevře a infekční zárodek, sporoplazma, z ní migruje krví přímo do té tkáně, v níž se pak množí a rostou sporotvorná plazmodia. Za jedinečnost myxosporidií mezi prvoky byla pokládána jejich mnohobuněčnost a s ní související funkční a tvarová diferenciace buněk. Přesto však byly myxosporidie stále počítány mezi prvoky.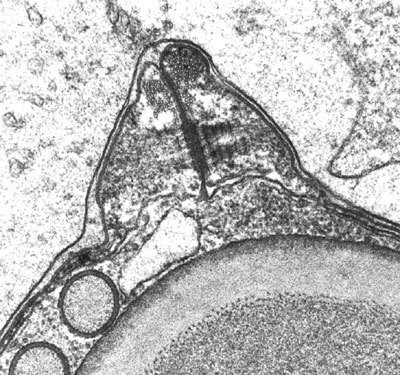 Tímto pojetím otřásl nástup elektronové mikroskopie. Potvrdila předchozí domněnky, že ve vývoji myxosporidií existuje zárodečná linie buněk, která přetrvává a z níž se neustále odvozují specializované somatické buňky, jako třeba velká mnohojaderná plazmodia nebo obalové buňky spor. Nedochází ovšem k tvorbě gamet v podobě spermií a oocytů. Překvapivý byl i nález desmozomů, tedy typických buněčných spojů metazoí, které spojují buňky tvořící se spory (viz obrázek). Desmozomy jsou jednou z charakteristik tkání živočichů. Nejzajímavější ale bylo zjištění, že pólové váčky mají nejen strukturu identickou s některými typy nematocyst, ale že se také identickým způsobem formují, mají stejnou morfogenezu. To je zcela odlišuje od poněkud podobných vymrštitelných organel jiných protistů, jako jsou třeba bičíkovci obrněnky rodu Polykrikos, kde však tyto organely vznikají naprosto odlišným způsobem.
Tímto pojetím otřásl nástup elektronové mikroskopie. Potvrdila předchozí domněnky, že ve vývoji myxosporidií existuje zárodečná linie buněk, která přetrvává a z níž se neustále odvozují specializované somatické buňky, jako třeba velká mnohojaderná plazmodia nebo obalové buňky spor. Nedochází ovšem k tvorbě gamet v podobě spermií a oocytů. Překvapivý byl i nález desmozomů, tedy typických buněčných spojů metazoí, které spojují buňky tvořící se spory (viz obrázek). Desmozomy jsou jednou z charakteristik tkání živočichů. Nejzajímavější ale bylo zjištění, že pólové váčky mají nejen strukturu identickou s některými typy nematocyst, ale že se také identickým způsobem formují, mají stejnou morfogenezu. To je zcela odlišuje od poněkud podobných vymrštitelných organel jiných protistů, jako jsou třeba bičíkovci obrněnky rodu Polykrikos, kde však tyto organely vznikají naprosto odlišným způsobem.
>To vše se klonilo ve prospěch názorů zejména Roberta Weilla, ředitele mořské biologické stanice ve Wimmereux ve Francii. Ten pokládal myxosporidie za potomky parazitizmem zjednodušených (neříkejme degenerovaných - jak víme, co z hlediska prospěšnosti v nějakém prostředí je pokrok?) vícebuněčných živočichů, patrně žahavců. Vedla ho k tomu zejména podobnost pólových váčků s nematocystami.
Vývoj v rybě tvoří jen polovinu života myxosporeí
Nečekaným obratem v poznání myxosporeí byla zpráva otištěná v Science (K. Wolf a M. E. Markiw, 1984), že vývoj probíhající v rybě tvoří pouhou polovinu jejich úplného životního cyklu. Ken Wolf se v západní Virginii v USA zabýval celý život rybími viry a stal se v tomto oboru klasikem. Pracoval většinou s viry pstruha duhového a tak mu neušlo, že vrtohlavost pstruhů, nebezpečné onemocnění na pstružích farmách, působené myxosporidií druhu Myxobolus cerebralis, se dá jen velice nesnadno experimentálně přenést na zdravé ryby, a to jen pokud spory údajně „zrály“ v bahně na dně. Pokusné přenosy myxosporeových nákaz se vůbec až dotud dařily jen výjimečně. Virolog Wolf tehdy učinil svůj první a poslední, ale zato zásadní exkurz do protozoologie. Přesným, mnohonásobně ověřeným experimentem prokázal, že zralé spory M. cerebralis uvolňované z těla nakažené ryby nejsou pro další ryby vůbec infekční, ale že naproti tomu jsou infekční pro nitěnky druhu Tubifex tubifex (obrázek).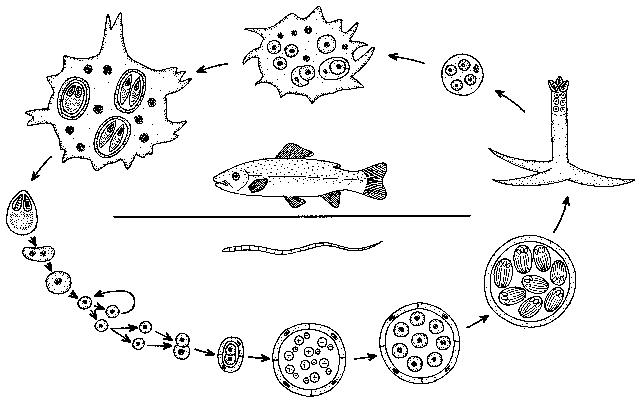 V nich se ze spor vyvine organizmus identický s prvokem rodu Triactinomyxon, který byl až dotud řazen do třídy Actinosporea.
V nich se ze spor vyvine organizmus identický s prvokem rodu Triactinomyxon, který byl až dotud řazen do třídy Actinosporea.
Organizmy této třídy objevil koncem minulého století v nitěnkách ve Vltavě český zoolog Štolc. Liší se od myxosporeí dlouhými plovákovitými cípy stěny spory, která obsahuje na rozdíl od myxosporidií velký počet infekčních zárodků. Je jich známo asi 40 druhů v několika rodech. Wolf objevil, že spory triactinomyxonu jsou neinfekční pro nitěnku, ale nakazí rybu, v níž se z nich opět vyvinou myxosporidie druhu Myxobolus cerebralis. Tím se uzavře vývojový cyklus, který svou složitostí nemá mezi protisty obdoby. Střídají se v něm dva samy o sobě úplné ale morfologicky odlišné cykly - myxosporidiový a aktinosporeový - zahrnující celkem dva typy vegetativního množení, dva odlišné sexuální procesy a dva typy sporogeneze končící tvorbou odlišných infekčních spor. A aby těm překvapením nebyl konec, ukázalo se, že zatímco nitěnka se skutečně nakazí pozřením spory, u ryb dojde k průniku parazita do těla většinou kůží, na kterou se přichytí spora aktinosporeové fáze z nitěnky.
Wolfovy výsledky se pochopitelně setkaly s nedůvěrou téměř všech protistologů. V čem by měl být smysl tak překomplikovaného životního cyklu? Tato otázka visí ve vzduchu stále. Teprve po třech letech se objevila první potvrzující práce a od té doby, během celých jedenácti let, bylo podobných přenosů mezi sladkovodní rybou a máloštětinatým červem experimentálně uskutečněno pouze patnáct. Je to počet jistě postačující k tomu, aby tento složitý cyklus byl pokládán za prokázaný i aby druhy dosud známé jako aktinosporidie byly chápány jen jako vývojová stadia myxosporeí. 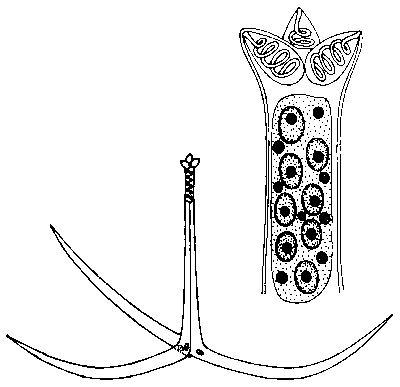 Nicméně omezený počet úspěšných přenosů, skutečnost, že přenosy se zdaleka ne vždy daří, i údaje o několika skutečně přímých přenosech z ryby na rybu, to vše ponechává místo k pochybnostem: Je tento složitý cyklus nutný ve všech případech? A platí i u velice hojných myxosporeí z mořských ryb, když z mořského prostředí jsou známy jen tři nálezy aktinosporidií? Zjištění, že různé druhy rodu Myxobolus, vzájemně si velmi podobné, mohou mít velmi odlišné aktinosporeové fáze, povede k úplnému přehodnocení dosavadní klasifikace myxosporidií. Mají se třídit podle toho, jak vypadá jejich fáze v rybě (jako dosud), nebo i podle utváření stadií v kroužkovcích?
Nicméně omezený počet úspěšných přenosů, skutečnost, že přenosy se zdaleka ne vždy daří, i údaje o několika skutečně přímých přenosech z ryby na rybu, to vše ponechává místo k pochybnostem: Je tento složitý cyklus nutný ve všech případech? A platí i u velice hojných myxosporeí z mořských ryb, když z mořského prostředí jsou známy jen tři nálezy aktinosporidií? Zjištění, že různé druhy rodu Myxobolus, vzájemně si velmi podobné, mohou mít velmi odlišné aktinosporeové fáze, povede k úplnému přehodnocení dosavadní klasifikace myxosporidií. Mají se třídit podle toho, jak vypadá jejich fáze v rybě (jako dosud), nebo i podle utváření stadií v kroužkovcích?
Etapa molekulární biologie je v historii výzkumu myxosporidií již třetím dramatickým zvratem
Srovnávání sekvence nukleotidů genu malé jednotky ribozomální nukleové kyseliny slouží jako obecně uznávaný metr na fylogenetické a příbuzenské vztahy všech organizmů. Jeho použití pořádně zpřeházelo přijaté příbuzenské vztahy uvnitř i mezi jednotlivými skupinami protistů. U myxosporeí byl dopad analýzy daleko radikálnější: tato skupina se vymyká z rámce říše protistů - nejsou příbuzní s žádnou z jejich skupin - a řadí se mezi metazoa, vícebuněčné živočichy. K těmto výsledkům dospěla shodně skupina amerických autorů v čele se Smothersem i práce, na níž se podílejí pracovníci Parazitologického ústavu AV ČR. Podivné přitom je, že příbuznost s láčkovci, předpokládaná pro strukturální shodu pólových váčků a nematocyst, se podle dosavadních výsledků nepotvrdila. Myxozoa se na dendrogramech řadí k bilateriím, tj. k živočichům dvoustranně souměrným, a nikoli k radialiím se souměrností paprsčitou, jako jsou žahavci (obrázek). Podle toho, jakou metodou jsou sekvence 18S rRNA srovnávány se sekvencemi ostatních eukaryot, řadí se myxozoa buď přímo mezi bilateria (metoda neighbor-joining), nebo jako sesterská skupina všech bilaterií (metoda maximum parsimony). Zvlášť podivné přitom je, že v obou případech jsou jim nejbližší skupinou červi hlístice! A tak dosavadní výsledky molekulárně biologického studia myxosporeí otevírají více otázek, než dávají odpovědí.
Podle toho, jakou metodou jsou sekvence 18S rRNA srovnávány se sekvencemi ostatních eukaryot, řadí se myxozoa buď přímo mezi bilateria (metoda neighbor-joining), nebo jako sesterská skupina všech bilaterií (metoda maximum parsimony). Zvlášť podivné přitom je, že v obou případech jsou jim nejbližší skupinou červi hlístice! A tak dosavadní výsledky molekulárně biologického studia myxosporeí otevírají více otázek, než dávají odpovědí.
Pokládáme-li myxozoa za parazitizmem degradované vícebuněčné živočichy, smíříme se i s tím, že podstoupily drastickou redukci své organizace, jedinečnou i mezi parazity vůbec. Jak ale porozumět výskytu pólových váčků, strukturou shodných s nematocystami, nejsou-li obě skupiny, myxozoa a žahavci, vůbec příbuzní? Pod dojmem převládajících teorií o endosymbiotickém vzniku buněčných organel vyslovil S. Shostak před dvěma lety domněnku, že intersticiální (vmezeřené) buňky žahavců, které dávají vznik nematocystám, se mohly vyvinout z endobionta, nějakého organizmu, který se zahnízdil v těle žahavce. Takový endobiont už mohl mít váčky podobné těm, co mají myxozoa, a tak by se dala celá věc vysvětlit. Poněvadž se však předpokládá, že už společní předkové žahavců měli nematocysty, a protože sekvenční analýza ukázala, že myxozoa jsou mladší než žahavci, celá tato domněnka padá a záhada trvá i nadále.
Co dál?
Rozhodně je třeba další práce, která by definitivně vyloučila příbuznost s dalšími skupinami žahavců, a pak by rozhodla, zda myxozoa jsou jen sesterskou skupinou bilaterií, nebo jejich skutečnou součástí. Tyto práce vnesou jasno i do taxonomie uvnitř kmene Myxozoa, která zatím zřejmě velmi málo obráží skutečné vztahy jednotlivých skupin. Mohou k tomu přispět i další molekulární informace, jako příbuznost genů kódujících například tubuliny a a b nebo minikolageny.V každém případě je fascinující, jaká překvapení stále ještě skrývá obrovská říše protistů, od nichž jsou odvozeny všechny vícebuněčné organizmy.
Literatura
Schlegel M., Lom J., Stechmann A., Bernhard D., Leipe D., Dyková I., Sogin M. (1995). Analysis of SSU rRNA of Myxidium lieberkuehni brings evidence that Myxozoa are Metazoa related with Bilateria. Sborník 2nd. European Congress of Protistology, Clermont-Ferrand, str. 85.Shostak S. (1993). A symbiogenetic theory for the origins of cnidocysts in Cnidaria. BioSystems 29, 49-58.
Smothers J. F., von Dohlen C. D., Smith Jr. L. H., Spall R. D. (l994). Molecular evidence that the myxozoan protists are metazoans. Science 265, 1179-1121.
Wolf K., Markiw M. E. (1984). Biology contravenes taxonomy in the Myxozoa: new discoveries show alternation of invertebrate and vertebrate hosts. Science 225, 1449-1452
O autorovi
Jiří Lom
Doporučujeme

Ničí ozon choleru? 

Jak se člověk stává biologem
