









Potíže s kořenem
Eukaryotické organismy přestavují produkt fascinujícího evolučního bujení, jehož historii zkoumá už několikátá generace biologů. Jedním z hlavních úkolů je zkonstruovat fylogenetický strom, který by co nejvěrněji odrážel evoluci jednotlivých linií eukaryot, jak se odvětvovaly od dávného předka až po moderní zástupce.
Vývoj v této oblasti nabírá na obrátkách. Za to vděčíme především prudkému nárůstu objemu sekvenčních dat (sekvencí celých genomů nebo alespoň transkriptomů) pro zástupce nejroztodivnějších eukaryotických skupin a také pokroku v metodách fylogenetických, či lépe řečeno fylogenomických analýz.
Současný pokus o vyjádření aktuální představy o stromu života eukaryot (obr. 1) se liší od schématu publikovaného ve Vesmíru před sedmi lety.1) V novém stromu již nefigurují dvě dříve předpokládané, ovšem vždy kontroverzní „superskupiny“ eukaryot, Chromista a Excavata.2) Opuštění skupiny Chromista je samo o sobě zajímavé téma související s posunem znalostí o evoluci plastidů, nás zde ale bude více zajímat „zamítnutí“ skupiny Excavata, protože se přímo týká pozice kořene eukaryotické fylogeneze.
{kind=link}
Hledá se společný předek
Kořen fylogenetického stromu je označení pro nejhlubší místo větvení, de facto společného předka celé skupiny. Znát pozici kořene je nezbytný předpoklad k dedukci předkovy pravděpodobné podoby.
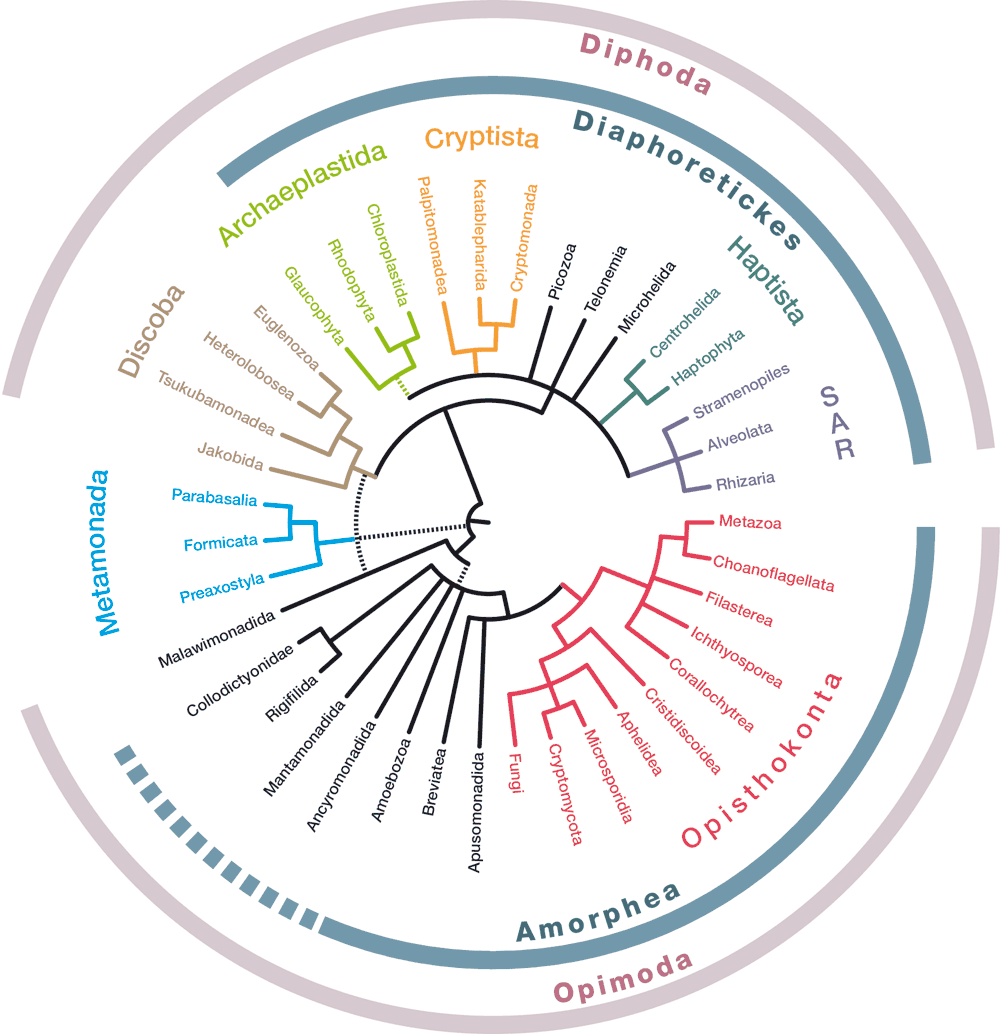 Seriózní snahy o určení nejhlubšího větvení ve fylogenetickém stromu eukaryot se datují přinejmenším do raných devadesátých let minulého století. V předchozím článku z roku 2010 jsme se pozice eukaryotického kořene stručně dotkli, ovšem se závěrem, že o ní nelze říci nic s jistotou. I dnes máme k jistotě daleko, ale vývoj byl natolik zajímavý a nadějný, že stojí za to podívat se na něj podrobněji.
Seriózní snahy o určení nejhlubšího větvení ve fylogenetickém stromu eukaryot se datují přinejmenším do raných devadesátých let minulého století. V předchozím článku z roku 2010 jsme se pozice eukaryotického kořene stručně dotkli, ovšem se závěrem, že o ní nelze říci nic s jistotou. I dnes máme k jistotě daleko, ale vývoj byl natolik zajímavý a nadějný, že stojí za to podívat se na něj podrobněji.
Jakým způsobem lze obecně stanovit pozici kořene fylogenetického stromu? Potřebujeme určit časovou posloupnost evolučních změn různých znaků studované skupiny organismů a pak hledat takovou pozici kořene (výchozího bodu celé fylogeneze), která je s touto posloupností v nejlepším souladu. Problém je v tom, že evoluce neupřednostňuje žádný směr nebo trend, takže v praxi je stanovení časové polarity evoluce znaků netriviální záležitostí.
Nejspolehlivějším řešením je použití „mimoskupiny“, v odborné hantýrce nazývané outgroup. Jde o organismy, o nichž máme dobrý důvod se domnívat, že fylogeneticky leží vně skupiny (označované jako ingroup), jejíž kořen hledáme. Outgroup poskytuje vodítko, jak evoluční změny jednotlivých znaků v rámci ingroup časově polarizovat. Znaky sdílené zástupci ingroup se zástupci outgroup jsou totiž pravděpodobně evolučně původní, zatímco možné odchylky u části zástupců ingroup jsou nejspíš pozdější evoluční změny, které se odehrály právě jen v určité podskupině naší ingroup. Kladení vajec ptakopyskem a ježurou, sdílené s plazí outgroup, je tak u savců podle všeho původní znak, zatímco rození živých mláďat je evoluční inovace definující odvozenost živorodých savců (a pozici kořene vně této skupiny).
Podobná logika se používá i při hodnocení molekulárních znaků, i když moderní fylogenetika se již nespoléhá na starou dobrou Occamovu břitvu a reálně pracuje s čím dál komplexnějšími statistickými přístupy.
První naděje… a zklamání
Takže zkusme určit pozici eukaryotické fylogeneze pomocí outgroup, což pochopitelně budou prokaryotické organismy. Budeme používat molekulární znaky, typicky sekvence homologických genů a proteinů, neboť jiné typy znaků pro takto dávné období evoluce nepřicházejí v úvahu.
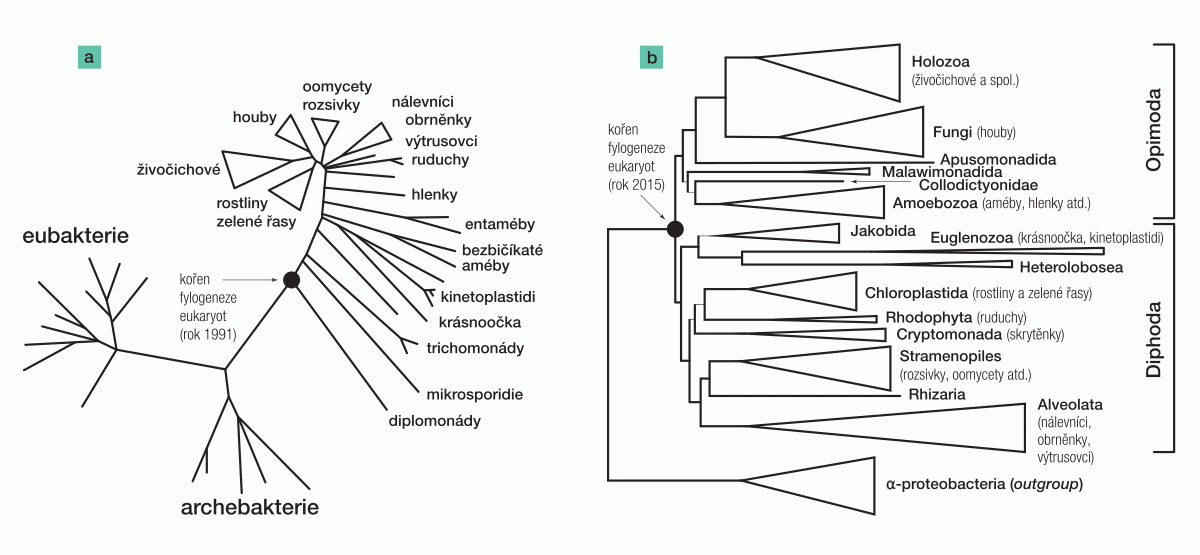
 První fylogenetické stromy zahrnující prokaryota i eukaryota se vykreslovaly už v počátcích molekulární fylogenetiky, především když americký biolog Carl Woese začal systematicky stanovovat a porovnávat sekvence „univerzálního fylogenetického markeru“, RNA složky malé podjednotky ribozomu (SSU rRNA). Tato molekula je přítomná u všech buněčných organismů a navíc je dostatečně konzervovaná, takže je možné ji smysluplně porovnávat i napříč eukaryoty a prokaryoty (díky tomu mohl Carl Woese mimo jiné rozpoznat archebakterie jako samostatnou doménu odlišnou od eubakterií). Již první „univerzální“ fylogenetické stromy na bázi sekvencí SSU rRNA publikované v osmdesátých letech 20. století tak vlastně automaticky ukazovaly konkrétní pozice kořene eukaryotické fylogeneze, my se ovšem zastavme až u stromu, který publikoval roku 1991 Mitch Sogin (obr. 2a).3)
První fylogenetické stromy zahrnující prokaryota i eukaryota se vykreslovaly už v počátcích molekulární fylogenetiky, především když americký biolog Carl Woese začal systematicky stanovovat a porovnávat sekvence „univerzálního fylogenetického markeru“, RNA složky malé podjednotky ribozomu (SSU rRNA). Tato molekula je přítomná u všech buněčných organismů a navíc je dostatečně konzervovaná, takže je možné ji smysluplně porovnávat i napříč eukaryoty a prokaryoty (díky tomu mohl Carl Woese mimo jiné rozpoznat archebakterie jako samostatnou doménu odlišnou od eubakterií). Již první „univerzální“ fylogenetické stromy na bázi sekvencí SSU rRNA publikované v osmdesátých letech 20. století tak vlastně automaticky ukazovaly konkrétní pozice kořene eukaryotické fylogeneze, my se ovšem zastavme až u stromu, který publikoval roku 1991 Mitch Sogin (obr. 2a).3)
{kind=link}
Tento dodnes často (obvykle nekriticky) přetiskovaný strom má charakteristickou strukturu: dlouhá větev oddělující eukaryota od prokaryot, série postupně se odvětvujících „bazálních“ eukaryotických linií a tzv. korunu zahrnující mnohobuněčná eukaryota spolu s některými protistními skupinami.
Kořen eukaryotické fylogeneze v tomto stromu z definice leží v místě, kde se k eukaryotickému „podstromu“ (subtree) připojuje větev směřující od prokaryot, tedy mezi skupinou zvanou diplomonády a ostatními eukaryoty. Následující hluboké eukaryotické větve pak představují mikrosporidie a trichomonády. Tyto tři skupiny na první pohled postrádají mitochondrie, takže topologie Soginova stromu podporovala hypotézu archezoí slavného evoluční protistologa Thomase Cavaliera-Smithe. Podle ní se většina hlavních vlastností eukaryotické buňky objevila ještě před endosymbiotickým vznikem mitochondrie a pre- -mitochondriální eukaryota, tedy Archezoa, dodnes žijí, reprezentovaná právě například diplomonádami.
Jakkoliv logicky to všechno zní, tento scénář vzal nakonec zasvé. Především se ukázalo, že domnělá dnešní Archezoa ve skutečnosti mitochondrie mají, i když ve značně pozměněné nebo redukované podobě.4) Paralelně vyšlo najevo, že topologie Soginova stromu je metodologický artefakt související se zrychlenou evolucí sekvencí genů „archezoí“, konkrétně tzv. long branch-attraction artefakt, při němž jsou rychle se vyvíjející eukaryotické linie „taženy“ k bázi eukaryotického stromu dlouhou větví separující prokaryota od eukaryot.
Podle dnešních znalostí všechna moderní eukaryota vzešla z předka obsahujícího mitochondrii a přinejmenším některé „archezoální“ skupiny zcela jistě nepředstavují hluboké eukaryotické linie (například mikrosporidie jsou prokazatelně blízce příbuzné houbám). Avšak ani dnes, kdy máme k dispozici dramaticky zlepšené metody konstrukce fylogenetických stromů a rozumíme vcelku dobře zdroji artefaktů ve fylogenetických analýzách, nejsme schopni spolehlivě určit pozici kořene eukaryotické fylogeneze na základě srovnávání sekvencí SSU rRNA genů. Na to v tomto jednom genu prostě není dost historického signálu.
S rozšiřující se databází genomových sekvencí se začalo experimentovat s dalšími geny, přesněji řečeno proteiny (sekvencemi aminokyselin), coby fylogenetickými markery pro rekonstrukci eukaryotické fylogeneze včetně určení pozice kořene. Jako nejvhodnější se zdály především dobře konzervované proteiny účastnící se základních procesů spojených s replikací genomu, genovou expresí a proteosyntézou (translací). Tuto mašinerii eukaryotická buňka zdědila po svých archebakteriálních předcích, proto by se dalo očekávat, že homologické sekvence dnešních archebakterií budou představovat vhodnou outgroup pro určení eukaryotického kořene.
Bohužel se ukázalo, že sekvence těchto markerů u eukaryot a u archebakterií (natož eubakterií) jsou příliš odlišné, zřejmě proto, že eukaryotická linie se oddělila od nejbližších známých archebakteriálních příbuzných již hodně dávno a její geny navíc asi prodělaly fázi zrychlené evoluce (lze si představit, že při „přestavbě“ archebakteriálních předků do podoby eukaryotické buňky se děly lecjaké dramatické evoluční události). Výsledkem jsou opět nevěrohodné výsledky zatížené artefakty a zatím není jasné, zda se nakonec sekvence archebakterií jako užitečnou outgroup pro eukaryota podaří použít.
Tápání ve tmě
Co zkusit pozici eukaryotického kořene určit nějak jinak? Ve zmíněném článku ve Vesmíru v roce 2010 jsme popsali další zajímavý pokus nám již známého Toma Cavalier-Smithe z prvních let nového tisíciletí. Tehdy se svými spolupracovníky přišel s představou, že kořen eukaryot leží mezi dvěma velkými eukaryotickými skupinami označenými jako Unikonta a Bikonta. Jak jsme ovšem mohli konstatovat již před sedmi lety, důvody pro vymezení těchto skupin, a tedy i pro kořen mezi nimi, se ukázaly jako fakticky neopodstatněné a tento závěr platí dodnes (přesto se dělení eukaryot na Unikonta a Bikonta bohužel stále objevuje v pracích nedostatečně informovaných autorů).
Cavalier-Smith ovšem jako neúnavný hledač neváhal přijít v roce 2010 s novým, radikálně odlišným nápadem, že totiž kořen eukaryotické fylogeneze leží mezi skupinou Euglenozoa a ostatními eukaryoty. Euglenozoa (zahrnující nejen vlastní krásnoočka – třeba rod Euglena, ale například i původce spavé nemoci Trypanosoma brucei) na molekulární úrovni skutečně vykazují některé neobvyklé rysy (např. jejich DNA obsahuje unikátně modifikované nukleotidové báze), to ale samo o sobě neznamená, že se musí jednat o linii sesterskou všem ostatním eukaryotům.
Hlavní argument se týkal proteinu Tom40, který je kritický pro import jaderně kódovaných proteinů do mitochondrie a je přítomen u všech eukaryot právě s výjimkou skupiny Euglenozoa. Zdálo se, že Tom40 u euglenzoí nemůže chybět druhotně jako důsledek ztráty, protože ztráta tohoto proteinu okamžitě znamená nefunkčnost mitochondrie. Proto Cavalier-Smith postuloval, že k oddělení evolučních linií euglenozoí a ostatních eukaryot došlo ještě před vznikem Tom40 a že Euglenozoa nezávisle vyvinula vlastní způsob, jak dostávat proteiny z cytoplazmy do mitochondrie. To se záhy zdálo být potvrzeno objevem proteinu ATOM u euglenozoí stojícího na místě Tom40. Jak ale ukázali Vojtěch Žárský a jeho spolupracovníci z Přírodovědecké fakulty UK v Praze, ATOM je ve skutečnosti divergentní homolog proteinu Tom40.5) Aktuálně tak neexistují věrohodné doklady pro bazální pozici euglenozoí v eukaryotické fylogenezi.
Jiní autoři použili jiný přístup k stanovení pozice eukaryotického kořene a přišli s dalšími návrhy. Skupina fenomenálního Eugena Koonina například v roce 2009 provedla analýzu tzv. vzácných genomových změn představovaných výjimečnými substitucemi nebo inzercemi či delecemi v sekvencích proteinů homologických napříč eukaryoty a prokaryoty (prokaryotické sekvence zde tedy představovaly outgroup). Tato analýza naznačila, že by eukaryotický kořen mohl ležet poblíž nebo dokonce uvnitř „superskupiny“ eukaryot s primárním plastidem (tzv. Archaeplastida). Tato studie ale trpěla velmi omezeným samplingem a otazníky panovaly i ohledně použité metodologie. (Je zajímavé, že autoři zatím neprovedli obdobnou analýzu s lepším samplingem, i když by tomu dnes nemělo nic bránit.)
Jiný přístup zvolil tým Laury Katzové, který v roce 2012 „spočítal“ fylogenetické stromy pro sérii eukaryotických genových rodin, a pak hledal takovou podobu zakořeněné eukaryotické fylogeneze, jež minimalizuje nutný počet genových duplikací a ztrát v evoluční historii analyzovaných genových rodin. Podle této analýzy by kořen měl ležet mezi „superskupinou“ Opisthokonta a ostatními eukaryoty, případně dokonce uvnitř této superskupiny. Jak nás ale učí zkušenost, takové parsimonní (na úspornost soustředěné) uvažování se s evoluční realitou obvykle míjí, nehledě na to, že některé aspekty fylogenetického stromu plynoucího z této analýzy jsou v přímém rozporu s dobře doloženými příbuzenskými vztahy eukaryot.
Mitochondrie jako klíč
Výše jsem poněkud skepticky mluvil o možném využití archebakterií jako dobré outgroup pro nalezení kořene eukaryotické fylogeneze, a to v důsledku příliš velké divergence mezi geny eukaryot a arachebakterií. Eukaryotická buňka však kromě genové výbavy zděděné od archebakteriálních předků obsahuje i výraznou eubakteriální komponentu – mitochondrii. Jak již jsem uvedl v souvislosti s pádem hypotézy Archezoa, dnes není pochyb o tom, že všechna moderní eukaryota vzešla z předka obdařeného mitochondrií. Různé nezávislé analýzy pak jednoznačně dokládají, že mitochondrie sama vznikla z endosymbiotické α-proteobakterie a že s tímto endosymbiontem (protomitochondrií) do eukaryotické buňky přišlo množství genů, z nichž většina dnes sedí v jaderném genomu (a jen nepatrný zbytek zůstává v genomech samotných mitochondrií). Podstatnou otázkou je, kdy k pohlcení protomitochondrie došlo. Jedná se o krajně kontroverzní téma, ale podle mého názoru se čím dál přesvědčivější jeví scénář, který počítá s relativně pozdním příchodem protomitochondrie do eukaryotické linie.6)
Pokud je tedy mitochondrie relativně pozdní evoluční inovace eukaryot, pak by distance mezi geny dnešních α-proteobakterií a geny vnesenými do eukaryot s protomitochondrií nemusela být tak velká, jako je divergence mezi geny dnešních archebakterií a „archebakteriálními geny“ v dnešních eukaryotech (přesně tohle dokazuje studie citovaná v pozn. 6)). Jinými slovy, geny moderních α-proteobakterií by potenciálně mohly být využity jako dobrá outgroup pro stanovení pozice kořene eukaryotické fylogeneze, samozřejmě za předpokladu, že „α-proteobakteriální geny“ v eukaryotech jsou vhodným markerem pro stanovení fylogenetických vztahů v rámci eukaryot samotných.
První test tohoto předpokladu učinili v roce 2012 mladý fylogenetik Romain Derelle a matador studia evoluce mitochondrií B. Franz Lang. Ti ukázali, že fylogenetické vztahy uvnitř eukaryot rekonstruované na základě eukaryotických genů α-proteobakteriálního původu (přesněji – proteinů kódovaných těmito geny) poskytují výsledky velmi dobře odpovídající jiným, častěji používaným markerům. Použití genů z dnešních α-proteobakterií jako outgroup pak naznačilo možnou pozici eukaryotického kořene, ovšem tato analýza trpěla nedostatečným samplingem (mnoho důležitých eukaryotických skupin bylo reprezentováno chabě nebo chybělo úplně) a nepřesvědčivými hodnotami statistické podpory získaných výsledků.
Tyto nedostatky se však brzy podařilo z větší části odstranit, a to i díky spolupráci Romaina Derella s mojí výzkumnou skupinou na Ostravské univerzitě a se skupinou Čestmíra Vlčka v Ústavu molekulární genetiky AV ČR v Praze. Výsledkem této spolupráce byl významně rozšířený sampling eukaryotické diverzity (díky našemu sekvenování genomů zástupců některých obskurních, avšak klíčových eukaryotických linií) a v dalších ohledech zlepšený fylogenomický dataset, konkrétně sada eukaryotických proteinových sekvencí kódovaných geny α-proteobakteriálního původu spolu s homologickými sekvencemi z moderních α-proteobakterií.
Analýzy tohoto datasetu za použití aktuálních metod fylogenomiky poskytly velmi silnou podporu pro místo, kde se větev vedoucí od α-proteobakteriání outgroup připojuje k eukaryotickému „podstromu“, tj. pro pozici kořene eukaryotické fylogeneze (obr. 2b).7) Tento kořen rozděluje eukaryota na dvě základní „megaskupiny“, pro něž Romain navrhl jména Opimoda a Diphoda (akronymy odvozené od názvů hlavních skupin, které do těchto megaskupin spadají). Na takto určené pozici kořene je nejpozoruhodnější fakt, že nabourává monofylii předpokládané superskupiny Excavata. Je to proto, že jedna linie exkavát, malawimonády, spadá do megaskupiny Opimoda, zatímco další linie exkavát, Discoba, spadá do megaskupiny Diphoda. Tento výsledek má ve skutečnosti zásadní dopad na rodící se představu o podobě buňky raných eukaryot včetně jejich předka, ale tohle vzrušující téma záměrně opomenu s příslibem, že se k němu v dohledné době na stránkách Vesmíru vrátím.
Konečně jasno?
Za výsledky naší práce si s Romainem Derellem samozřejmě stojíme, ale v životě, vědě a fylogenetice obzvlášť není nikdy nic jistého. Námi navrženou pozici eukaryotického kořene proto bude nutné podrobit dalším testům a nejlepší by bylo, kdyby se ji podařilo potvrdit nějakou analýzou nezávislou na α-proteobakteriálních genech.
Něco takového se vlastně už zčásti podařilo. Ještě před publikováním našeho článku vyšla obdobně zaměřená studie konkurenční skupiny Sandry Baldaufové z Uppsaly.8) Autoři této analýzy také vyšli z úvahy, že eubakteriální komponenta v genomech eukaryot může posloužit k určení pozice eukaryotického kořene, ovšem nesoustředili se na geny z protomitochondrie, ale využili jakékoliv eubakteriální geny, které se do eukaryot dostaly (zřejmě procesem horizontálního přenosu) ještě před existencí jejich posledního společného předka. Takových genů z jiných skupin než α-proteobakterie je překvapivé množství, což ve skutečnosti značí, že moderní eukaryotická buňka vznikla integrací mnoha prvků (nejen vlastního archebakteriálního základu s protomitochondriálním endosymbiontem).
Fylogenomické analýzy těchto genů s využitím eubakteriální outgroup vedly k závěru, že kořen eukaryotické fylogeneze leží mezi skupinou Discoba a ostatními eukaryoty, tedy jinde, než navrhuje naše práce. Tato studie však měla podstatně horší sampling než ta naše a Romain navíc pečlivou kontrolou použitých dat ukázal, že u některých genů byly namísto těch správných sekvencí použity odlišné varianty genů vzniklé dávnými duplikacemi. Stručně: byla porovnávána jablka s hruškami (pro fajnšmekry – paralogy namísto ortologů). Po odstranění tohoto problému dává „eubakteriální“ dataset (geny volené Sandrou) stejný výsledek ohledně pozice eukaryotického kořene jako náš „α-proteobakteriální“ dataset. Nejde v tomto případě o zcela nezávislé analýzy, protože oba datasety část genů sdílejí, ovšem shoda výsledků je povzbudivá.
I pokud se naše hypotéza o pozici kořene bude dále potvrzovat, je ve skutečnosti nekompletní. Pozice některých eukaryotických skupin zatím nemohla být testována, protože pro ně nebyla k dispozici odpovídající genomová data. Máme však dobré důvody domnívat se, že většina těchto dosud ignorovaných linií bude jednoznačně patřit mezi Opimoda nebo mezi Diphoda.
Největší otazník se vznáší nad pozicí skupiny Metamonada, která zahrnuje většinu skupin dříve považovaných za Archezoa, například diplomonády nebo trichomonády (viz výše). Metamonády nemohly být do našich analýz zahrnuty, protože ve svých genomech obsahují minimum genů α-proteobakteriáního původu. Důvodem je, že většina těchto genů je funkčně spojena s klasickou aerobní mitochondrií, takže se ztrácejí, pokud se mitochondrie mění na anaerobní organelu (hydrogenozom nebo mitozom). S metamonádami je ovšem ještě další problém. Fylogenomické analýzy zatím nedokázaly spolehlivě určit jejich pozici ani v nezakořeněných stromech (bez prokaryotické outgroup) s použitím obvyklých fylogenetických markerů. V závislosti na použitých genech, výběru taxonů a zvolených fylogenetických metodách metamonády vycházejí buď jako sesterská linie skupiny Discoba (pak by patřily mezi Diphoda), nebo jako sesterská linie malawimonád (pak by patřily mezi Opimoda), nebo jako samostatná větev někde mezi Opimoda a Diphoda (jak je tomu v naší nedávno publikované studii9)).
Poslední případ připouští možnost, že eukaryotický kořen leží mezi metamonádami a ostatními eukaryoty – skoro jako by se dveřmi vyhnaná hypotéza Archezoa chtěla oknem vrátit zpět. No, uvidíme. V každém případě si na definitivní určení pozice eukaryotického kořene budeme muset ještě počkat.
Poznámky
1) Vesmír 89, 464, 2010/7
2) Do Excavata se řadily například trichomonády, krásnoočka nebo Naegleria fowleri, améba způsobující smrtelnou meningoencefalitidu (příběh české epidemie z šedesátých let: Vesmír 93, 489, 2014/9). Mezi Chromista patřili například nálevníci, Toxoplasma, Plasmodium (původce malárie), obrněnky, skrytěnky, rozsivky nebo dírkonošci.
3) Sogin M. L., Curr. Opin. Genet. Dev., DOI: 10.1016/S0959-437X(05)80192-3
4) Podrobněji viz Vesmír 95, 354, 2016/6.
5) Zarsky V. et al., Current Biology, DOI: 10.1016/j.cub.2012.03.057
6) Viz například pozoruhodná studie Pittis A. A., Gabaldón T., Nature, DOI: 10.1038/nature16941
7) Derelle R. et al., PNAS, DOI: 10.1073/pnas.1420657112
8) He D. et al., Current Biology, DOI: 10.1016/j.cub.2014.01.036
9) Karnkowska A. et al., Current Biology, DOI: 10.1016/j.cub.2016.03.053
Ke stažení
 článek ve formátu pdf [417,72 kB]
článek ve formátu pdf [417,72 kB]
O autorovi
Marek Eliáš

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
