










O přirozenosti společenstev: ptáci a krajina
Ekologie se tradičně rozděluje na autekologii (studium vztahu jednotlivých druhů k prostředí), ekologii populací, ekologii společenstev a ekologii ekosystémů. Na rozdíl od ostatních podoborů není v případě ekologie společenstev zcela jasně vyhraněn předmět studia. Záleží totiž na tom, jak společenstvo chápeme – zda jako víceméně uzavřený a opakovatelný soubor organizmů různých druhů, jakýsi superorganizmus, který vzniká a udržuje se na základě výhodné kooperace mezi jeho členy, nebo jako soubor organizmů, které se vyskytují na jednom místě pouze na základě podobných nároků na prostředí.
 Kontroverze mezi těmito dvěma koncepcemi je charakteristická zejména pro ekologii rostlinných společenstev, zatímco v ekologii živočišných a konkrétně ptačích společenstev bylo vždy poměrně zřejmé, že oba extrémy jsou značně zjednodušující. Předmět studia ekologie společenstev tak podle mého názoru nejlépe vystihuje podivuhodně „nevědecká“ MacArthurova definice společenstva: „Community is any set of organisms currently living near each other and about which is interesting to talk“, tedy „jakékoli seskupení organizmů žijících blízko sebe, které stojí za řeč“! Dodejme, že v ekologii společenstev (přinejmenším živočišných) není větší autorita než MacArthur.
Kontroverze mezi těmito dvěma koncepcemi je charakteristická zejména pro ekologii rostlinných společenstev, zatímco v ekologii živočišných a konkrétně ptačích společenstev bylo vždy poměrně zřejmé, že oba extrémy jsou značně zjednodušující. Předmět studia ekologie společenstev tak podle mého názoru nejlépe vystihuje podivuhodně „nevědecká“ MacArthurova definice společenstva: „Community is any set of organisms currently living near each other and about which is interesting to talk“, tedy „jakékoli seskupení organizmů žijících blízko sebe, které stojí za řeč“! Dodejme, že v ekologii společenstev (přinejmenším živočišných) není větší autorita než MacArthur.
{kind=link}
Klasický náhled na společenstva
Ekologie společenstev se zrodila z tušení, že v přírodě je nějaký řád. Stačí přijít do listnatého lesa a dívat se, kde všude sbírají ptáci potravu. Zjistíme jednak, že každý druh ji sbírá jinde a jiným způsobem, jednak, že mezi jednotlivé druhy jsou všechna potenciální místa jaksi rozdělena: na zemi sbírají drozdovití ptáci, pod kůrou kmenů brhlík a šoupálci a hlouběji datlovití, z listů sbírají hmyz pěnice a budníčci, sýkory obírají větvičky a lejsci zase loví hmyz poletující ve vzduchu. Ekologie společenstev se snaží hledat zdroj tohoto tušeného řádu. Tradičně je zdroj jakéhokoli řádu spatřován v nějakém omezení. Ekologové se proto obrátili k témuž principu jako Darwin: totiž k malthusovské představě růstu populace. Podle této představy platí, že za předpokladu stabilního a homogenního prostředí budou populace růst až do okamžiku, kdy se zdroje stanou limitními a natalita dosáhne rovnováhy s mortalitou. V tomto okamžiku bude prostředí saturováno, zdroje budou využívány beze zbytku a dvě populace si nutně musí začít konkurovat (=kompetovat), poněvadž se vzájemně připravují o zdroje. Konkurenčně schopnější populace vzápětí vytlačí druhou populaci – dojde ke konkurenčnímu vyloučení (obrázek). Mezidruhová konkurence je v rámci této představy nutným důsledkem populačního růstu a (vzhledem k negativnímu účinku) nutným důsledkem této konkurence je zase selekční tlak na její omezení – tedy na rozdělení zdrojů mezi dva druhy. Důsledkem společného využívání zdrojů je ekologická separace druhů, zajišťující koexistenci. „Řád přírody“ – jak rozdílnou prostorovou distribuci, tak rozdílné využívání zdrojů v rámci jednoho společenstva jednotlivými druhy – můžeme proto vysvětlovat na základě přirozeného výběru.Potud vše v pořádku. Vzhledem k tomu, že mezidruhová konkurence a její důsledky vyplývají – jak jsme si ukázali – nevyhnutelně z teorie, ekologie se dlouhou dobu nemusela zabývat konkurencí jako takovou (ta je přece nutná), ale způsoby koexistence druhů – jak je možné, že dva druhy se mohou společně vyskytovat na jednom místě. Rozdíly mezi druhy byly tedy vysvětlovány ze společenstva, respektive z koevoluce potenciálně konkurujících druhů. Problém nastal, když se začaly zkoumat předpoklady zmíněné teorie, totiž ona homogenita a stabilita. Zjistilo se, že alespoň v suchozemském prostředí tyto předpoklady v podstatě nikdy nejsou splněny, že vždy (na každé měřítkové úrovni) je prostředí heterogenní, mozaikovité, a navíc proměnlivé. Co to znamená pro představu konkurence jako faktoru určujícího diverzifikaci mezi druhy?
Společenstvo v mozaikovitém a proměnlivém prostředí
Za předpokladu heterogenity prostředí se začne jevit role konkurence v poněkud jiném světle. Mohou nastat následující případy:- Druhy si nemusí konkurovat z těchto příčin:
- Populace nedorostou až na úroveň, kde je zdroj limitní, neboť jejich početnost je snižována jiným způsobem než limitujícími zdroji (např. disturbancemi).
- I drobná odlišnost druhů vede ke koexistenci, která není vyvolána selekčním tlakem (každý druh si najde místo, které je pro něj výhodnější).
- Každý druh čte environmentální mozaiku v jiném měřítku: např. sýkora koňadra rozlišuje les – neles a na typu lesa už jí nezáleží, zatímco třeba králíček obecný vyhledává striktně jen les jehličnatý. Dva druhy, které jsou tímto ve vztahu generalista – specialista, si nekonkurují, poněvadž se nemohou vzájemně připravovat o zdroj.
- Druhy si mohou konkurovat, ale bez selekčního efektu – konkurence je v mozaikovitém a proměnlivém prostředí téměř vždy jen lokální a krátkodobá a nemá vliv na dlouhodobé charakteristiky populace.
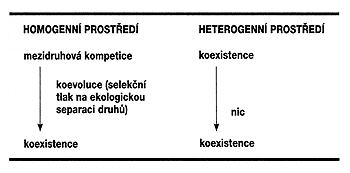 Vidíme tedy, že mozaikovitost a proměnlivost prostředí je rozhodující pro procesy v něm probíhající. V takovém prostředí se nemůže mezidruhová konkurence příliš uplatnit a druhy vždy spíše koexistují. Otázkou je, zda má vůbec smysl mluvit o koexistenci, když vlastně není jiná možnost, když nejde o výsledek nějakého procesu, ale o pouhé konstatování, že druhy žijí vedle sebe (obrázek). V tom případě by však nemělo smysl ani moc mluvit o společenstvech, když – viz MacArthurovu definici – to, že druhy žijí vedle sebe, „nestojí za řeč“. Důsledkem zpochybnění společenstev jako seskupení koexistujících druhů je individualistické pojetí ekologie, představa, že druhy jsou vzájemně nezávislé a že hledání onoho řádu v přírodě je scestné.
Vidíme tedy, že mozaikovitost a proměnlivost prostředí je rozhodující pro procesy v něm probíhající. V takovém prostředí se nemůže mezidruhová konkurence příliš uplatnit a druhy vždy spíše koexistují. Otázkou je, zda má vůbec smysl mluvit o koexistenci, když vlastně není jiná možnost, když nejde o výsledek nějakého procesu, ale o pouhé konstatování, že druhy žijí vedle sebe (obrázek). V tom případě by však nemělo smysl ani moc mluvit o společenstvech, když – viz MacArthurovu definici – to, že druhy žijí vedle sebe, „nestojí za řeč“. Důsledkem zpochybnění společenstev jako seskupení koexistujících druhů je individualistické pojetí ekologie, představa, že druhy jsou vzájemně nezávislé a že hledání onoho řádu v přírodě je scestné.
{kind=link}
Jsou druhy opravdu vzájemně nezávislé?
Zajímavé poučení nám v tomto ohledu skýtá diskuse o náhodnosti či nenáhodnosti osídlení ostrovů v jihozápadním Pacifiku. Americký ekolog J. M. Diamond sestavil jistá pravidla platná pro distribuce ptáků na ostrovech v okolí Nové Guiney (viz text v rámečku), na jejichž základě dokazoval vzájemnou závislost druhů. Na to ostře reagovali vědci Connor a Simberloff, kteří tvrdili, že tato pravidla vyplývají i z nulové hypotézy – tj. z předpokladu náhodného a nezávislého osídlení ostrovů. Nemíním zde rozebírat celou 11 let trvající diskusi. Podstatné je, že Connorova a Simberloffova nulová hypotéza se ukázala být ne tak úplně nulovou. Dokonale nulová hypotéza by totiž předpokládala, že na každém ostrově bude stejná pravděpodobnost výskytu daného druhu, ať už je ostrov jakkoli malý a ať už je daný druh jakkoli vzácný, což je zřejmý nesmysl. Aby vyloučili tyto zjevně nesmyslné možnosti (např. že na malém ostrově budou všechny druhy), sestavovali tuto hypotézu s následujícími omezeními:- celkový počet druhů na jednotlivých ostrovech odpovídá pozorovanému počtu druhů,
- každý druh se vyskytuje jen na ostrovech té velikostní třídy, která odpovídá jeho pozorovanému výskytu,
- každý druh osídluje takový počet ostrovů, který odpovídá pozorovanému počtu. Ukázalo se přitom, že již tato omezení v sobě vlastně zahrnují vzájemnou závislost druhů (např. to, že některé druhy se vyskytují jen na malých, druhově chudých ostrovech, není asi nic náhodného).
Z toho ovšem vyplývá jedna – možná triviální, přesto však podle mého názoru podstatná – skutečnost: ať už jde o projevy konkurence nebo jiných procesů, druhy jsou závislé už proto, že se nějak vztahují ke společnému prostředí. Druhy jsou tedy závislé už společným čtením prostředí (třeba tím, že dva druhy mají v oblibě tentýž typ ostrova, či naopak právě opačný typ ostrova). Otázkou ovšem zůstává, do jaké míry má toto čtení nějakou logiku, nějaký řád, či zda se vztahy k prostředí u jednotlivých druhů vyvíjely nezávisle a např. dva druhy žijí na jednom místě jen proto, že čtou podobně prostředí. To, že z takzvané nulové hypotézy lze vyvodit tak sugestivní pravidla (viz text v rámečku), svědčí ovšem spíše pro to, že nějaký řád by se za různými způsoby čtení mohl skrývat.
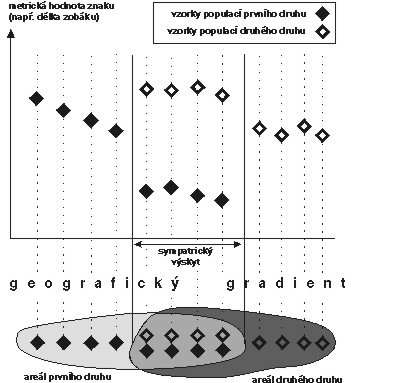
I některé další důkazy svědčí pro představu konkurence a následné koevoluce druhů, tedy pro to, že vztah k prostředí se vyvíjel u jednotlivých druhů ve vzájemné závislosti. Připomeňme si např. „character displacement“ (obrázek), tj. případ, kdy dva druhy jsou morfologicky odlišnější na místech, kde žijí sympatricky – tzn. kde se jedinci obou druhů mohou setkávat – než na místech, kde žijí alopatricky (každý zvlášť). Jiným příkladem je třeba mezidruhová teritorialita, která se někdy vyskytuje u příbuzných druhů ptáků. V obou případech jde o aktivní koexistenční mechanizmy, o koexistenci, která nespočívá jen v prostém společném výskytu – „character displacement“ morfologickým“ rozrůzněním přímo vylučuje potenciální konkurenci, mezidruhová teritorialita vylučuje nepříznivé důsledky této konkurence (stejným způsobem jako vnitrodruhová teritorialita vylučuje nepříznivé důsledky vnitrodruhové konkurence). Jak se tyto věci slučují s tím, že – jak bylo ukázáno – v heterogenním prostředí nemá klasická představa mezidruhové konkurence velkou šanci? 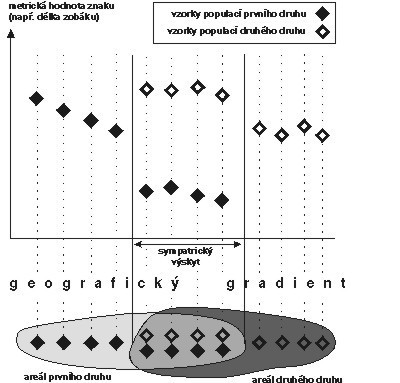
{kind=link}
Jak se může mezidruhová konkurence uplatnit v heterogenním prostředí?
Proti sobě tedy stojí dvě koncepce: individualistické pojetí společenstev a představa, že koexistence druhů ve společenstvech je produktem mezidruhové konkurence a následného přirozeného výběru. Z této kontroverze bychom, podle mého názoru, mohli najít východisko, když si uvědomíme následující momenty:- Heterogenita zároveň znamená nerovnocennost různých míst. Díky tomu se zdroje, které nejsou limitní, limitními stanou a mezidruhová konkurence se může uplatnit, přestože na poněkud jiné úrovni. Příkladem jsou sýkory koňadra a modřinka, které soutěží o hnízdní dutiny i v případě, že těchto dutin (budek) je několikanásobně více než hnízdících párů. Není totiž dutina jako dutina, některé jsou oblíbenější, některé méně a konkurence mezi těmito druhy probíhá formou šarvátek o oblíbené dutiny. V dobách vysoké populační hustoty sýkory koňadry jsou tak modřinky vytlačeny do méně oblíbených dutin. Jiným příkladem je konkurence mezi hmyzožravými ptáky: hmyzu může být sice dost, ale jen někde jej lze efektivně lovit a jen někde jej lze najít. Nezáleží tedy na jakémsi objektivním množství zdroje, ale na tom, jak jsou zdroje jednotlivými druhy rozlišovány (jako v případě oněch dutin), nebo na tom, jaké jsou vůbec možnosti jejich úspěšného vyhledání a zužitkování danými druhy. Pro možnost konkurence je tedy rozhodující čtení prostředí jednotlivými druhy, jak se jednotlivé druhy k prostředí vztahují, jak rozlišují.
- Co se týče mezidruhové konkurence, je nepochybné, že někde a někdy probíhá, existují pro to vcelku nesporné důkazy. Problémem zůstává, jaký je její dosah, když právě probíhá jen někde a někdy – jak na základě této velice lokální záležitosti (vzpomeňme na zmíněné sýkory) může časem dojít třeba k rovnoměrnému rozdělení zdrojů mezi dva druhy. Domnívám se, že i zde nám může pomoci představa nerovnocennosti míst v mozaikovitém prostředí a z ní plynoucí koncepce metapopulační dynamiky. Nerovnocennost míst se totiž může projevit i jejich nestejným významem pro přežívání a vývoj populací.
Koncepce metapopulační dynamiky říká v zásadě toto: žádná populace nemůže růst ve všech místech rovnoměrně. Pokud je celková populační hustota stabilní, znamená to, že v některých místech natalita převyšuje mortalitu (to jsou tzv. zdrojová místa) a v některých místech je tomu naopak. Ze zdrojových míst migrují jedinci do těch horších míst. Zdrojová místa (rep. procesy v nich probíhající) jsou tedy pro budoucnost populace důležitější než jiná. Potom ovšem může mít i lokální konkurence vliv na celkové charakteristiky populace, pokud bude probíhat právě v těchto zdrojových místech.
V případě, že právě ve zdrojových místech bude konkurence nejvýznamnějším selekčním faktorem, její vliv se projeví časem v celé populaci. Druhy tedy mohou být závislé, v jejich čtení prostředí je možno hledat nějaký řád, mohly se u nich (přinejmenším teoreticky) vyvinout mechanizmy oné aktivní koexistence. Procesy, které k nim vedou, však mohou mít velice jemnou povahu.
Lze tedy shrnout, že heterogenita, nerovnocennost jednotlivých míst, tak jak je čtena jednotlivými druhy, může být podstatná pro vznik onoho „řádu“, pro vznik aktivní koexistence. Většina procesů má velice lokální povahu, ale díky nerovnocennosti míst může mít mnohem globálnější dosah.
Alternativní možnosti konstituce společenstev
Dotkli jsme se skutečnosti, že schopnost organizmů vyhledat nějaký zdroj či předvídat příznivost nějakého místa je omezená. Zatímco v homogenním prostředí by to bylo lhostejné (když je všeho všude stejně, není potřeba nic hledat či předvídat), v heterogenním prostředí jsou na této schopnosti všechny organizmy závislé. Tato skutečnost se podle mého názoru může také významně podílet na konstituci společenstev. Jedním z nejlepších zdrojů informace o kvalitě daného místa může být totiž zkušenost příslušníka stejného druhu. Bylo zjištěno, že ptáci reagují pozitivně na výskyt příslušníků vlastního druhu, konkrétně že mnohé druhy hnízdí přednostně na těch místech, kde již zahnízdil jedinec téhož druhu (i když nejde o koloniální, ale naopak o teritoriální druh). Tento dosud poměrně opomíjený efekt konspecifického přitahování má možná pro strukturu společenstev větší význam, než jsme kdy tušili. Lze si navíc představit podobný efekt i v rámci třeba skupiny příbuzných druhů. Výskyt jiných druhů může být stejně hodnotným ukazatelem kvality prostředí, jako třeba typ vegetace. Čtení prostředí se tedy nemusí omezovat jen na environmentální mozaiku, ale i na blízce příbuzné organizmy a tento efekt se může projevit na struktuře společenstev. Omezení jednotlivých organizmů z hlediska jejich schopnosti vnímat důležité parametry prostředí může v heterogenním prostředí být důležité i pro to, jak se budou organizmy číst navzájem, a tedy i pro to, jak se budou konstituovat společenstva.Vidíme tedy, že onen tušený „řád přírody“ může vznikat na mnohem subtilnější a jemnější úrovni, než bychom byli předpokládali – druhy si mohou konkurovat jen díky podobnému rozlišování vhodných a nevhodných míst, konkurence je vždy lokální, ale její dosah je závislý na povaze místa, kde probíhá, a v heterogenním prostředí má rozhodující význam čtení prostředí i vzájemné čtení druhů. Heterogenita tedy není jen přitěžující komplikace, ale je naopak základní, neredukovatelnou vlastností přírody, díky níž mají ekologické procesy poněkud jinou, jemnější, složitější, ale možná zajímavější logiku. Ukazuje se totiž, že rozhodující procesy mohou mít sice v podstatě mechanickou povahu, ale podmínky těchto procesů, tedy to, co určuje, jak a kdy budou probíhat, jsou veskrze biologické, ne-li psychologické povahy – záleží na vlastnostech a biologických omezeních jednotlivých druhů, na tom, jak druhy čtou prostředí, jak rozlišují, co považují za podstatné a co za nepodstatné. „Mechanická“ a biologicko-psychologická povaha procesů tak tvoří dva vzájemně komplementární póly, mezi nimiž probíhá dění v přírodě.
Vzhledem k tomu, co bylo řečeno o povaze prostředí a procesů v něm probíhajících, mám čím dál silnější dojem, že matricí, na které probíhají ekologické procesy, nejsou jakési více či méně uzavřené celky (ekosystémy), ale že touto matricí je spíše to, co jsme si zvykli označovat slovem krajina.
Pravidla platná pro distribuci ptačích druhů na ostrovech v okolí Nové Guiney:
- Příbuzné druhy se vyskytují společně jen v některých z možných kombinací.
- Tyto „dovolené“ kombinace jsou odolné proti invazi těch druhů, díky nimž by se staly „zakázanými“ kombinacemi.
- Kombinace, která je stabilní na velkém, druhově bohatém ostrově, může být nestabilní na malém, druhově chudém ostrově.
- Druhová kombinace na druhově chudém, malém ostrově, může být odolná vůči invazi těch druhů, které se snadno začlení do kombinace na velkém, druhově bohatém ostrově.
- Některé páry druhů se nikdy nevyskytují společně.
- Některé samy o sobě nestabilní páry druhů mohou být součástí větší, stabilní kombinace.
- Některé kombinace mohou být naopak nestabilní, přestože se skládají pouze z kombinací, které jsou samy o sobě stabilní.
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
