










Velikost genitálií indikuje klimatické změny
Pro rekonstrukci klimatu ve čtvrtohorách se v zásadě uplatňují dva přístupy. První – nazvěme jej třeba analytický – spočívá v precizním určení obsahu izotopů kyslíku ve vápencích či kadmia ve schránkách mořských měkkýšů nebo v nějaké podobné analytické metodě. Číslo je zde „pánembohem“, metody jsou velmi vědecké, drahé a důmyslné. Pokud se někde něco přehlédne, tak je to většinou chyba zásadního významu – třeba že schází část zkoumaného souvrství. Nicméně byl to právě tento analytický přístup, který paleoklimatologii postavil na pevné nohy reprodukovatelných řad čísel odehrávajících se na určité časové škále.
Druhý přístup nazvěme třeba přírodovědeckým. Nepracuje s obsahy prvků či jejich izotopů, ale s živými tvory, přesněji řečeno se zbytky toho, co kdysi živými tvory bývalo. Přírodovědné metody nebývají tak exaktní (ostatně ani život si nepotrpí na přesnost a vypočítatelnost), bývají lacinější a mají svá jiná interpretační úskalí. Klasickou přírodovědnou metodou je např. pylová analýza, při které v jednotlivých vrstvách sedimentů počítám pylová zrna a podle toho usuzuji např. na proměny lesního prostředí. Ukazuje se, že ve starších pracích býval silně nadhodnocen rozsah borů, protože borovice produkují obrovská množství pylu, který je vzdušnými proudy transportován na stovky kilometrů daleko, a navíc se dobře uchovává. Jinou oblíbenou, ale těžko interpretovanou veličinou je doba prvního rozkvětu nějaké rostliny. Ten je řízen celkovou klimatickou odezvou – nejenom teplotou, ale také vlhkostí či přítomností nočních mrazů a jen obtížně jej převedeme do grafu, který by vypadal tak exaktně jako např. graf obsahů stabilních izotopů. Nicméně trampové dobře vědí, že když kvetou tařice, tak je v noci ještě zima, když kvetou petrklíče, tak se to dá vydržet, a když přiletí vlašťovky, tak je možné vzít na výlet i malé děti.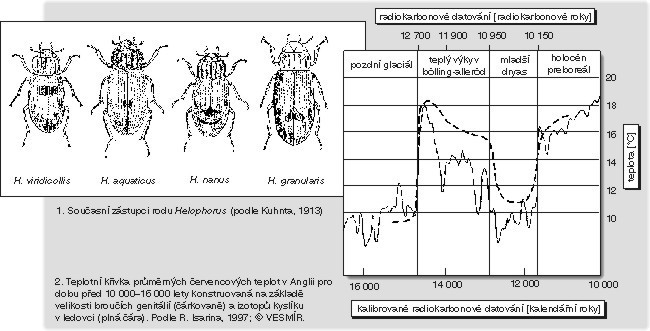
K běžným metodám patří určování typu prostředí pomocí společenstev fosilních měkkýšů a drobných obratlovců, zejména hlodavců. Obě skupiny jsou citlivé na kolísání vlhkosti a na míru zapojení či rozvolnění lesa. Hlodavci vzhledem k své větší mobilitě zachycují proměny prostředí v měřítku několika hektarů či km2, některé druhy měkkýšů však reagují na proměny stanoviště (např. rozdíl mezi severním a jižním úpatím téže skály) již v měřítku několika metrů. Obsahuje-li určitá vrstva více druhů fosilního záznamu – uhlíky stromů, pylová zrna, měkkýše, kosti obratlovců, popřípadě i keramické zlomky různých pravěkých kultur a to vše je uloženo v nějakém geologicky definovaném sedimentu, získáváme paralelní záznam různých lokálních, regionálních a kontinentálních událostí. Obvykle se potřebujeme opřít o globální vzorec klimatických změn, na které pak „zavěšujeme“ jemnější klimatické a environmentální odezvy např. lesního ekosystému střední Evropy až po lokální vývoj krajiny.
Jednou z novějších metod studia klimatických změn během několika posledních ledových dob je výzkum fosilní etnofauny. Ukazuje se, že zbytky vnější kostry mnoha desítek druhů brouků se sice rychle rozpadají, ale vydrží v sedimentech desítky tisíc let. Brouci navíc vnášejí do studia klimatických změn nový žádoucí rozměr: jsou nesmírně pohybliví, takže nový areál o rozměru třeba části kontinentu mohou obsadit již během několika let. Myším trvá podobná migrace desítky či stovky let a měkkýšům ještě déle (a to i když se vozí na ptácích). Fosilní i žijící brouci – Coleoptera – jsou v mnoha případech velmi citliví nejen na prostředí, ale také na průměrné teploty v době rozmnožování. V sedimentech někdy nalézáme vedle sebe různé druhy brouků, z nichž některé jsou citlivé na jarní a jiné na letní a podzimní průměrné teploty. Některé druhy brouků jsou vázány na padlé stromy, jiné na určitou skladbu dřevin. Při dobré znalosti paleoekologie je tedy možné z druhové skladby brouků vyčíst řadu údajů. Vypadá to krásně, ale přírodovědcům jsou okamžitě jasné problémy této metody.
Řada druhů hmyzu má cyklický průběh četnosti, a to možná i bez ohledu na klimatické změny. Většina cyklů souvisí s životním cyklem populace a trvá jako u chrousta několik let, ale pravděpodobně mohou existovat i delší cykly četnosti, které trvají nejméně desítky let. Další problém je určení druhů. Z rozpadlé kostry fosilního hmyzu se totiž spolehlivě dají určit jen genitálie, jejichž velikost je obvykle menší než 1 mm. Pro paleoklimatická studia je často používán rod Helophorus, pro jehož jednotlivé druhy je charakteristická velikost genitálií. Ta je hlavním rozlišovacím znakem mezi chladnomilnými sibiřskými druhy a teplomilnějšími druhy středních šířek.
Na následujícím grafu můžete srovnat dvě paleoklimatické křivky. Ta spodní, plná a detailní, je konstruována na kombinovaném základě objemu ledu v ledovcích a izotopových hodnotách pro kyslík v ledovci. Ta horní, čárkovaná a méně přesná, je konstruována na základě velikosti genitálií brouka rodu Helophorus. Obě jsou vztaženy na Britské ostrovy a na průměrnou červencovou teplotu. Kromě viditelné zásadní shody zbývá vysvětlit rozdíly v obou křivkách. Především teplota získaná na základě velikosti broučích genitálií je pravděpodobně přesnější než izotopová teplota, která je založena na modelu. Na druhou stranu je možné fosilní brouky vyplavit jen ze suchozemských sedimentů, které nemají tak dobrou časovou rozlišovací schopnost jako roční přírůstky v ledovcích a krátké změny prostě nezachytí. Důležité je, že izotopovou křivku je možné kalibrovat pomocí „broučí“ křivky, co se týče skutečné teploty, a pak se spolehnout na detailní ledovcový záznam, takže nakonec si fyzik a entomolog mohou potřást rukama.
Kromě toho si všimněte další důležité věci – podle tabulky začíná preborál, jenž je prvním obdobím holocénu, asi před 11 700 lety, a nikoliv před 10 000 lety, jak bylo vždy udáváno. Onen rozdíl necelých 2000 let je přitom velmi důležitý, protože se do něj vejde důležitá část mezolitického vývoje lidstva. Dřívější datum bylo založeno na radiokarbonovém datování, zatímco dnešní datování je založeno na ročních přírůstkových liniích ledovců a jezerních sedimentů. Jak je možné, že se ta data podstatně rozcházejí? Na sklonku poslední ledové doby se totiž radiokarbonové hodiny na dobu téměř jednoho tisíce let zastavily. Tím, jak se koncem ledové doby rozbíhala oceánická cirkulace, vynášely hluboké mořské proudy obrovská množství oxidu uhličitého, který ředil CO2 v atmosféře. Podobný jev, jemuž říkáme Suessův efekt, známe i v průmyslové době, kdy je atmosférický CO2 ředěn antropogenním oxidem uhličitým uvolňovaným spalováním fosilních paliv. Výsledek je ten, že na poslední dvě století funguje radiokarbonové datování velmi špatně, což nám nevadí, protože si můžeme pomoci jinými metodami, např. metody s využitím izotopů olova, ale podstatně nám to překáží v jednom z nejdůležitějších období vývoje lidstva – na konci poslední ledové doby, kde místo jednoduchého radiokarbonu musíme trpělivě (a často zbytečně) počítat přírůstkové linie.
Citát
R. C. Lewontin: Biologie jako ideologie, Jota, Brno 1997, s. 74
Vzdělané a horní vrstvy často vzhlížejí k jiným kulturám a hledají u nich to nejlepší. Vědci z anglicky mluvících zemí jsou často zváni k rozhovorům do italské televize a jejich odpovědi v angličtině jsou simultánně překládány do italštiny. Když jsou autoři těchto programů dotazováni, proč nepřizvou k debatě italské vědce, odpovídají, že Italové jednoduše nevěří vědeckým poznatkům formulovaným italsky a že se přesvědčí pouze tehdy, slyší-li daná tvrzení pronesená v jazyce anglickém.Ke stažení
 Článek ve formátu PDF [433,4 kB]
Článek ve formátu PDF [433,4 kB]
O autorovi
Václav Cílek

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
