










Molekulové motory
Jednou z nápadných a také podstatných vlastností živých organizmů je aktivní pohyb, kdy se uvnitř organizmu přetváří některá z forem energie v energii pohybovou. (Ne tedy pád těla z pátého poschodí, ale naopak výstup po schodech.) Aktivní pohyb organizmů (tzv. lokomoce) vůči okolí pomocí svalů, bičíku (u spermií) či pohyb améboidní (u měňavek) je jen jednou z nejnápadnějších forem biologického pohybu. Jiný aktivní pohyb, ne vůči okolí, najdeme uvnitř mnohobuněčných živočichů: stahuje se srdce, „pracují“ střeva, mluvíme, dýcháme. To vše jsou ovšem také pohyby svalové. A jestliže se podíváme na život jedné buňky, zjistíme, že i on je spojen s neustálým vnitřním pohybem: cytoplazma uvnitř buňky proudí, buněčné organely se organizovaně přemísťují z místa na místo, pomocí pohybu měchýřků buňka přijímá látky z okolí (pinocytóza), či pohlcuje větší částečky (fagocytóza), vydává látky (exocytóza), ve vláknech nervových buněk neustále proudí látky k nervovým zakončením a směrem opačným, při dělení buňky putují chromozomy do dceřiných buněk a buňka se aktivně zaškrcuje. Tedy kdekoliv je život, setkáváme se s aktivními pohyby nejrůznějších forem, na nejrůznějších úrovních, nejrůznějšího smyslu, účelu a cíle, jak se dočteme v každé učebnici biologie. Nové je jen to, že všechny zmíněné formy pohybu spojuje stejný princip – funkce molekulových motorů. K tomu dospěla věda až v posledních deseti letech, a proto pojem molekulový motor najdeme jen v moderních učebnicích.
Molekulový motor je oligomerní protein s enzymovou aktivitou
Motorem rozumíme obecně nějaký stroj či zařízení, které přeměňuje některou z forem energie v energii kinetickou. Ve spalovacím motoru se přeměňuje chemická energie, v elektromotoru elektrická, v motorech biologické povahy energie chemická. Ta se uvolní nejčastěji při hydrolytickém rozštěpení – z adenozintrifosforečné kyseliny (ATP). Ta je tedy „palivem“. A jak molekulový motor vypadá? Je to komplex několika molekul bílkovin (oligomerní protein). Z toho vyplývá i molekulová hmotnost „motorů“ a jejich rozměr. Kdybychom si je představili jako kuličky (viz obrázek), pak by jejich průměr byl asi 20 nm (1 nm = 1 miliontina milimetru). Kdybychom je řadili do „šňůrky“, vešlo by se jich do jednoho milimetru 50 000. Za to, že člověk může přeskočit překážku vyšší než dva metry, jsou primárně odpovědné tak malinké struktury. Nestačí k tomu ovšem jediný molekulový motor, musí jich synchronně pracovat miliony.
A jak molekulový motor vypadá? Je to komplex několika molekul bílkovin (oligomerní protein). Z toho vyplývá i molekulová hmotnost „motorů“ a jejich rozměr. Kdybychom si je představili jako kuličky (viz obrázek), pak by jejich průměr byl asi 20 nm (1 nm = 1 miliontina milimetru). Kdybychom je řadili do „šňůrky“, vešlo by se jich do jednoho milimetru 50 000. Za to, že člověk může přeskočit překážku vyšší než dva metry, jsou primárně odpovědné tak malinké struktury. Nestačí k tomu ovšem jediný molekulový motor, musí jich synchronně pracovat miliony.
Jaká je podstata funkce molekulových motorů, či mechanizmus transformace chemické energie v energii mechanickou? Motory mají vlastní enzymovou aktivitu, tj. katalyzují hydrolýzu adenozintrifosforečné kyseliny (jsou to tedy ATPázy). Při rozštěpení makroergní vazby ATP se uvolní energie, která změní tvar motoru, a to tak, že se v prostoru poněkud posune po nějaké podložce. Při hydrolýze další molekuly ATP se motor opět posune stejným směrem. Při opakování hydrolytických cyklů se pak motor po podložce stále pohybuje po určité dráze (jako auto po silnici či vlak po kolejích). Vzdálenost, o kterou se motor posune po podložce při jednom cyklu, je kolem 10 – 20 nm. Rychlost, kterou se motory pohybují, je pozoruhodná, až 100 nm za sekundu, takže dráhu 1 mm by motor urazil za 10 sekund (tj. 360 mm/hod). Rychlost pohybu většiny motorů je však menší, při neuronovém transportu, kdy se dopravují neurotransmitery k nervovým zakončením, to je 0,5 – 4 m/sec, tj. 400 mm za den.
 Logika strukturní organizace je u všech motorů stejná (viz obrázek a obrázek). Všechny mají tzv. motorovou doménu, která má enzymovou aktivitu (štěpí ATP) a současně se posunuje po podložce. Je zajímavé, že polypeptidický řetěz motorové domény má velmi konzervativní strukturu (velké podobnosti v sekvenci aminokyselin), i když jde o motory, které se v evoluci vyvíjely patrně nezávisle na sobě. Na motorovou doménu je připojeno několik polypeptidů dalších (s různou funkcí) a na ně koncový polypeptid (koncová doména), který má specifickou afinitu k některým jiným molekulám či buněčným strukturám. Z hlediska pohybové aktivity jako takové je tedy důležitá motorová doména, z hlediska specifické pohybové funkce v buňce koncová doména.
Logika strukturní organizace je u všech motorů stejná (viz obrázek a obrázek). Všechny mají tzv. motorovou doménu, která má enzymovou aktivitu (štěpí ATP) a současně se posunuje po podložce. Je zajímavé, že polypeptidický řetěz motorové domény má velmi konzervativní strukturu (velké podobnosti v sekvenci aminokyselin), i když jde o motory, které se v evoluci vyvíjely patrně nezávisle na sobě. Na motorovou doménu je připojeno několik polypeptidů dalších (s různou funkcí) a na ně koncový polypeptid (koncová doména), který má specifickou afinitu k některým jiným molekulám či buněčným strukturám. Z hlediska pohybové aktivity jako takové je tedy důležitá motorová doména, z hlediska specifické pohybové funkce v buňce koncová doména.
Motory se pohybují po strukturách cytoskeletu
Neméně důležitá pro pohyb je podložka, po níž se motory posunují. Jsou to výlučně struktury cytoskeletu, konkrétně mikrotubuly a mikrofilamenta (viz Vesmír 65, 678, 1986/12). Mikrotubuly jsou tenké trubičky, mají 25 nm v průměru a jsou tvořeny bílkovinou tubulinem. Mikrofilamenta jsou tenká vlákna (7 nm v průměru), tvořená bílkovinou aktinem (obrázek). Tyto trubičky a vlákna najdeme v cytoplazmě všech buněk (s výjimkou buněk prokaryontních) a ve všech buněčných strukturách specializovaných na pohybové funkce, jako jsou bičíky, řasinky, pseudopodie, dělicí vřeténka, myofibrily svalových buněk ad. Ve zmíněném článku ve Vesmíru bylo uvedeno, že jednou z hlavních funkcí cytoskeletu je podíl na transformaci chemické energie v mechanickou. Jak tedy souvisejí molekulové motory s cytoskeletem?
Mikrotubuly jsou tenké trubičky, mají 25 nm v průměru a jsou tvořeny bílkovinou tubulinem. Mikrofilamenta jsou tenká vlákna (7 nm v průměru), tvořená bílkovinou aktinem (obrázek). Tyto trubičky a vlákna najdeme v cytoplazmě všech buněk (s výjimkou buněk prokaryontních) a ve všech buněčných strukturách specializovaných na pohybové funkce, jako jsou bičíky, řasinky, pseudopodie, dělicí vřeténka, myofibrily svalových buněk ad. Ve zmíněném článku ve Vesmíru bylo uvedeno, že jednou z hlavních funkcí cytoskeletu je podíl na transformaci chemické energie v mechanickou. Jak tedy souvisejí molekulové motory s cytoskeletem?
Hydrolýzu ATP spustí motor jedině kontaktem s mikrotubulem, resp. mikrofilamentem. Motory jsou tedy ATPázy aktivované tubulinem, resp. aktinem. Zároveň je mikrotubulus či mikrofilamentum onou pevnou podložkou, po které se motor pohybuje. Podobně se v metru kontaktem vozu s kolejnicí napájenou elektrickou energií spustí hnací agregáty (motory), vůz jede po kolejnici, která zároveň vymezuje pohyb vozu v prostoru. Přeruší-li se kontakt s trolejem (nebo dodávka proudu do kolejnice), vůz se zastaví. Dráha vozu je určena kolejnicí a lze ji měnit jedině tak, že se koleje postaví jinam. Jedině tak lze také měnit „dráhu“ molekulových motorů.
Simulační pokusy
Funkci molekulových motorů lze simulovat v pokusech in vitro (mimo buňku). Není obtížné připravit izolované mikrotubuly, izolovat motory jako bílkoviny v čistém stavu a izolovat ATP.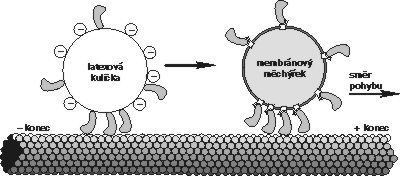 Motory jsou příliš malé na to, aby jejich pohyb bylo možno pozorovat např. ve světelném mikroskopu, lze je však zviditelnit tím, že se na ně navážou nějaké větší částečky, např. latexové kuličky (viz obrázek). Máme-li v systému pouze mikrotubuly a latexem značené motory, k žádnému pohybu nedochází. Jakmile se přidá „palivo“ (ATP), pohyb započne, a zastaví se až když „dojde benzin“ (ATP se vyčerpá). V simulačních pokusech lze zjistit, zda je pohyb motoru vázán na mikrotubuly, či na mikrofilamenta, jaká je rychlost jeho pohybu, energetická účinnost transformace ad.
Motory jsou příliš malé na to, aby jejich pohyb bylo možno pozorovat např. ve světelném mikroskopu, lze je však zviditelnit tím, že se na ně navážou nějaké větší částečky, např. latexové kuličky (viz obrázek). Máme-li v systému pouze mikrotubuly a latexem značené motory, k žádnému pohybu nedochází. Jakmile se přidá „palivo“ (ATP), pohyb započne, a zastaví se až když „dojde benzin“ (ATP se vyčerpá). V simulačních pokusech lze zjistit, zda je pohyb motoru vázán na mikrotubuly, či na mikrofilamenta, jaká je rychlost jeho pohybu, energetická účinnost transformace ad.
Typy molekulových motorů
Některé motory jsou aktivovány tubulinem a pohybují se podél mikrotubulů, jiné jsou aktivovány aktinem a pohybují se podle mikrofilament. Víme dnes, že oba systémy, mikrotubulární i aktinový, vznikly (evolučně) na sobě nezávisle. Svědčí pro to charakteristické různosti ve stavbě bílkovinných podjednotek nemotorové domény, ač motorové domény jsou velmi podobné. Motory vázané na mikrofilamenta (aktin) se obecně nazývají myoziny (patří sem i dlouho známý myozin svalových buněk). Motory vázané na mikrotubuly se obecně nazývají kineziny (z řeckého kinesis = pohyb), nebo dyneiny (z řeckého dynamis = síla). Proč rozlišení na kineziny a dyneiny? Vraťme se k metru. Vlak se po jedné koleji a jednom troleji může pohybovat tím nebo opačným směrem, ke změně směru stačí zařízení v hnacím agregátu (koleje nejsou polarizovány). To patrně molekulové motory nedovedou, pohybují se buď tím, nebo opačným směrem (jako kdyby pro každý směr musela být jiná lokomotiva). Příčina je v tom, že mikrotubuly i mikrofilamenta polarizovány jsou. Jejich konce se liší dynamikou připojování nepolymerizovaného tubulinu, resp. aktinu. Jeden konec se konvenčně označuje plus-konec, druhý je minus-konec. Kineziny se pohybují od plus-konce mikrotubulů k jejich minus-konci, dyneiny od minus-konce k plus-konci (viz obrázek). Všechny dosud známé myoziny se pohybují od minus-konce mikrofilament k jejich plus-konci.
To patrně molekulové motory nedovedou, pohybují se buď tím, nebo opačným směrem (jako kdyby pro každý směr musela být jiná lokomotiva). Příčina je v tom, že mikrotubuly i mikrofilamenta polarizovány jsou. Jejich konce se liší dynamikou připojování nepolymerizovaného tubulinu, resp. aktinu. Jeden konec se konvenčně označuje plus-konec, druhý je minus-konec. Kineziny se pohybují od plus-konce mikrotubulů k jejich minus-konci, dyneiny od minus-konce k plus-konci (viz obrázek). Všechny dosud známé myoziny se pohybují od minus-konce mikrofilament k jejich plus-konci.
Složitější pohybové funkce motorů
Lokomotiva se může pohybovat po kolejích, ale o to nejde, její smysl je v tom, aby něco vozila. Stejně tak pohyb molekulového motoru po příslušných drahách je jen elementární funkcí, která slouží nějakému účelu. Buď jsou motory volné a pohybují se v prostoru po mikrotubulu, který je k něčemu upevněn, nebo jsou samy někde upevněny, a pak pohybují mikrotubulem. V tom případě jsou motory vázány nejčastěji na jiný mikrotubulus a musí pak posouvat mikrotubuly vůči sobě. (Asi tak, jako když jdu po prkně, které je na válečcích, a nesu prkno nad hlavou: Které z prken se pohybuje vůči okolí? Evidentně to, které není na nic fixováno. Jedno vůči druhému se pohybují v každém případě.)Při prvním mechanizmu motory fungují jako nákladní automobil či vlak, tj. přenášejí (převážejí) něco někam, při druhém posouvají mikrotubuly či mikrofilamenta vůči sobě (jako kdyby vlak metra posouval koleje vůči nástupišti). První mechanizmus se uplatňuje zřetelně při transportu uvnitř buňky, druhý např. při pohybu řasinek a při pohybu svalovém.
Vnitrobuněčný transport
Příkladem prvního mechanizmu, kdy motory fungují jako přenašeče (translokátory), je vnitrobuněčný transport. Mohou přenášet něco někam či odněkud, pokud tam vedou dráhy (mikrotubuly či mikrofilamenta). Mikrotubuly jsou v buňce velmi přesně polarizovány. Jejich minus-konec je vázán na centrozom, který leží v blízkosti jádra, a jejich volné konce směřují k periferii buňky (viz obrázek). Kineziny tedy přemísťují „náklad“ (např. exocytózové měchýřky vznikající z Golgiho aparátu) od středu buňky k periferii a dyneiny naopak od povrchu buňky k jejich středu (např. endocytární měchýřky či fagocytární vakuoly). V buňce se neustále přemísťuje množství látek a struktur z jednoho místa na druhé, a to cíleně, podle potřeb buňky. V tomto smyslu je analogie mezi buněčnými dráhami a železnicí přesná, v něčem však přece pokulhává. Většinu železničních tratí máme z dob Marie Terezie (a stále slouží), kdežto v buňce se dráhy tvořené mikrotubuly a mikrofilamenty přestavují každou sekundu. Celá navigace v rozvozu materiálu je tedy dána neustálou přestavbou cytoskeletárních komponent. Představy o tom, jak je tato přestavba a tím celý vnitrobuněčný transport řízen, jsou zatím velmi mlhavé. I řada dalších otázek je zatím nejasná. Když jede nákladní vlak z Prahy do Brna, musí někdo vědět, co se má na vagony naložit, tj. co se má do Brna dovézt. Translokátory jsou specifické podle cytoskeletární struktury a podle směru, nikoliv však podle toho, co „vezou“. Musí tedy v buňce být nějaký mechanizmus, který to „ví“. Naše představy o buněčných „dopravních dispečerech“ se již začínají rýsovat. Jsou to opět molekuly bílkovin, označují se jako adaptory. Zaměřeny jsou jednak na přenášenou částečku (náklad), jednak na určitý typ přenašeče.
Jejich minus-konec je vázán na centrozom, který leží v blízkosti jádra, a jejich volné konce směřují k periferii buňky (viz obrázek). Kineziny tedy přemísťují „náklad“ (např. exocytózové měchýřky vznikající z Golgiho aparátu) od středu buňky k periferii a dyneiny naopak od povrchu buňky k jejich středu (např. endocytární měchýřky či fagocytární vakuoly). V buňce se neustále přemísťuje množství látek a struktur z jednoho místa na druhé, a to cíleně, podle potřeb buňky. V tomto smyslu je analogie mezi buněčnými dráhami a železnicí přesná, v něčem však přece pokulhává. Většinu železničních tratí máme z dob Marie Terezie (a stále slouží), kdežto v buňce se dráhy tvořené mikrotubuly a mikrofilamenty přestavují každou sekundu. Celá navigace v rozvozu materiálu je tedy dána neustálou přestavbou cytoskeletárních komponent. Představy o tom, jak je tato přestavba a tím celý vnitrobuněčný transport řízen, jsou zatím velmi mlhavé. I řada dalších otázek je zatím nejasná. Když jede nákladní vlak z Prahy do Brna, musí někdo vědět, co se má na vagony naložit, tj. co se má do Brna dovézt. Translokátory jsou specifické podle cytoskeletární struktury a podle směru, nikoliv však podle toho, co „vezou“. Musí tedy v buňce být nějaký mechanizmus, který to „ví“. Naše představy o buněčných „dopravních dispečerech“ se již začínají rýsovat. Jsou to opět molekuly bílkovin, označují se jako adaptory. Zaměřeny jsou jednak na přenášenou částečku (náklad), jednak na určitý typ přenašeče.
Analogie mezi železniční dopravou a buněčným transportem má však ještě jedno slabé místo. Vlaky se vyloží a na zpáteční cestu znovu naloží zbožím – recyklují. Molekulární motory se však mohou pohybovat jen jedním směrem, ať něco vezou, nebo ne. Jejich recyklace zatím není prokázána. Pro buňku by to asi bylo neeknomické, ale jak to buňka řeší, to zatím nevíme. Snad to naznačuje nález jednoho kinezinu (označovaného jako ncd-kinezin). Tento kinezin se jako všechny kineziny pohybuje od minus-konce k plus-konci mikrotubulů. Je-li však jeden z jeho lehkých řetězců fosforylován, začne se pohybovat opačným směrem. Nejnověji byl nalezen další kinezin tohoto typu (Kar3-kinezin).
Kdy používá buňka pro vnitřní transport mikrotubulární a kdy aktinový systém? Zajímavé je, že to patrně není univerzální. Např. pro exocytózu, tj. pro transport exocytózových měchýřků od Golgiho aparátu k periferii buňky, používá většina buněk kineziny, jiné používají myoziny (např. buňky hub). Některé mají dokonce naprogramovány oba systémy a při alternativním vyřazení jednoho z nich stačí k zajištění exocytózy ten zbývající.
Klouzání mikrotubulů a mikrofilament
Druhý mechanizmus pohybových buněčných funkcí uskutečňovaný molekulovými motory je posun cytoskeletárních komponent, tj. mikrotubulů podél mikrotubulů a mikrofilament vůči mikrofilamentům (označuje se jako klouzáni mikrotubulů či mikrofilament). Přiblížíme stručně pouze dva pohybové procesy: pohyb kinocilií a pohyb svalový.Bičíky a řasinky (obecně kinocilie) jsou poměrně složité struktury. Jejich hlavní pohybovou součástí je centrální osové vlákno neboli axonema (ř. axon – osa, nema – vlákno). Je tvořena pravidelně rozestavěnými mikrotubuly: v ose řasinky jsou umístěny dva mikrotubuly a kolem nich je rozestaveno devět zdvojených mikrotubulů (dublet). Toto uspořádání se označuje jako „vzorec 9 + 2“ a ten je shodný u všech eukaryontních organizmů, od prvoků až po člověka.  Mezi dubletami jsou (mimo jiné) umístěna tzv. dyneinová ramena. Jsou tvořena molekulovým motorem – axonemálním dyneinem (viz obrázek). Tento dynein se poněkud liší od cytoplazmatického dyneinu – má více polypeptidových řetězců a tím i složitější strukturu. Váže se pevně svou koncovou doménou na jednu dubletu a volně – svými motorovými doménami – na dubletu sousední. Kdybychom řasinku či bičík od buňky odstřihli a přidali ATP, pak bude dynein posouvat mikrotubuly (dublety) navzájem a délka kinocilie by se prodloužila v podstatě devětkrát (9 dublet). Lze to elegantně experimentálně prokázat. V neporušených řasinkách či bičících jsou však dublety svými konci zakotveny v tzv. bazálním tělísku (jsou jedním koncem upevněny). Jestliže se hydrolyzuje ATP, je vzájemný posun mikrotubulů omezený, produkovaná energie ale dokáže celé osové vlákno ohnout. Logickou pravidelnou regulací aktivity dyneinů jednak mezi jednotlivými dubletami a jednak podél centrálního osového vlákna dochází ke složitým pohybům bičíku a řasinek v prostoru.
Mezi dubletami jsou (mimo jiné) umístěna tzv. dyneinová ramena. Jsou tvořena molekulovým motorem – axonemálním dyneinem (viz obrázek). Tento dynein se poněkud liší od cytoplazmatického dyneinu – má více polypeptidových řetězců a tím i složitější strukturu. Váže se pevně svou koncovou doménou na jednu dubletu a volně – svými motorovými doménami – na dubletu sousední. Kdybychom řasinku či bičík od buňky odstřihli a přidali ATP, pak bude dynein posouvat mikrotubuly (dublety) navzájem a délka kinocilie by se prodloužila v podstatě devětkrát (9 dublet). Lze to elegantně experimentálně prokázat. V neporušených řasinkách či bičících jsou však dublety svými konci zakotveny v tzv. bazálním tělísku (jsou jedním koncem upevněny). Jestliže se hydrolyzuje ATP, je vzájemný posun mikrotubulů omezený, produkovaná energie ale dokáže celé osové vlákno ohnout. Logickou pravidelnou regulací aktivity dyneinů jednak mezi jednotlivými dubletami a jednak podél centrálního osového vlákna dochází ke složitým pohybům bičíku a řasinek v prostoru.
Podstatou svalového pohybu je vzájemný posun mikrofilament, „poháněný“ molekulovým motorem myozinem. Podobnost s klouzáním mikrotubulů je vysoká.  Svalový myozin má však jednu zvláštnost: ve svalových buňkách se spojuje ve vlákna (viz obrázek). Myozinové vlákno je tvořeno tisíci myozinových molekul.
Svalový myozin má však jednu zvláštnost: ve svalových buňkách se spojuje ve vlákna (viz obrázek). Myozinové vlákno je tvořeno tisíci myozinových molekul.  V myofibrile, která je vlastním pohybovým aparátem svalové buňky, leží pak aktinová mikrofilamenta a myozinová vlákna paralelně vedle sebe (viz obrázek). Při hydrolýze ATP se aktinová a myozinová vlákna navzájem posunují a tím způsobí kontrakci složitěji organizované struktury (myofibrily). V jednom vláknu myozinu je mnoho motorových domén, a jejich energetický efekt, tj. množství transformované chemické energie v energii kinetickou, se sčítá. V buňkách příčně pruhovaných, srdečních a hladkých svalů je organizace aktinových a myozinových vláken jiná. (Nejsložitější je u příčně pruhovaného svalstva.) V každém případě je svalový pohyb – ve své energetické dokonalosti –
V myofibrile, která je vlastním pohybovým aparátem svalové buňky, leží pak aktinová mikrofilamenta a myozinová vlákna paralelně vedle sebe (viz obrázek). Při hydrolýze ATP se aktinová a myozinová vlákna navzájem posunují a tím způsobí kontrakci složitěji organizované struktury (myofibrily). V jednom vláknu myozinu je mnoho motorových domén, a jejich energetický efekt, tj. množství transformované chemické energie v energii kinetickou, se sčítá. V buňkách příčně pruhovaných, srdečních a hladkých svalů je organizace aktinových a myozinových vláken jiná. (Nejsložitější je u příčně pruhovaného svalstva.) V každém případě je svalový pohyb – ve své energetické dokonalosti –  jen evoluční obdobou molekulárního mechanizmu, tj. pohybu molekulového motoru (myozinu) po aktinovém mikrofilamentu (viz obrázek). Evolučně primitivní „svalíčky“ najdeme také bez výjimky v každé eukaryontní buňce. Uskutečňují proudění cytoplazmy, améboidní pohyb, fagocytózu, zaškrcení buňky při dělení atp.
jen evoluční obdobou molekulárního mechanizmu, tj. pohybu molekulového motoru (myozinu) po aktinovém mikrofilamentu (viz obrázek). Evolučně primitivní „svalíčky“ najdeme také bez výjimky v každé eukaryontní buňce. Uskutečňují proudění cytoplazmy, améboidní pohyb, fagocytózu, zaškrcení buňky při dělení atp.
Radost z poznání obecných principů
Kolotání cytoplazmy v buňce zelené řasy a kolotání lidských párů v tanečním reji má z hlediska elementárních energetických transformací stejný princip. Je to udivující? Pro laika možná, pro biologa již méně. Biologové totiž vědí, že podstata živých organizmů, ať se od sebe jakkoliv liší, je stejná a že stejné jsou i principy základních životních funkcí. Je fascinující tyto principy poznávat, ale je to složité a zdlouhavé. Vždyť myozin (ze svalových buněk) objevil R. Kuhn již r. 1864, aktin B. Straub r. 1942, axonemální dynein r. 1964 a jednoduchý cytoplazmatický myozin Pollard a Korn r. 1970. Na společný jmenovatel se toto vše převedlo tedy poměrně nedávno. Na otázky, které si K. Purkyně kladl před mnoha lety, bylo proto možno odpovědět až nyní. Nádherně se však potvrdila jedna věta z jeho práce o řasinkovém pohybu z r. 1834: „Zjev totiž, který se jeví v souhrnu ústrojů, na jedné straně není určen pro žádnou určitou funkci, na druhé straně zase je složkou všech funkcí, poněvadž neslouží k jednotlivým oněm úkonům, vloženým v samotné ústroje, ale jakožto věc všeobecná a základní tvoří základ nebo princip všech.“ 1)Poznámky
Citát
A BŮH STVOŘIL INTERNET
[...] Obecně je možno říci, že směrovací tabulky (na jejich základě se v síti rozesílají jednotlivé pakety – „balíčky“ – informací, pozn. red.) se aktualizují vždy, kdykoliv se změní konfigurace sítě; jinak by nebylo možné posílat pakety vždy tou nejkratší a nejjistější cestou. V sedmdesátých letech, kdy vznikaly první verze Arpanetu, by „normálním“ řešením bylo konstruovat tyto tabulky počítačem v „administrativním centru“, které by hlídalo stav všech linek a všech připojených počítačů. Na základě toho by toto centrum propočítalo tabulku nejvýhodnějších cest a tu by pak distribuovalo na všechny směrovače. Avšak tvůrci Arpanetu, později Internetu, vynaložili velkou námahu, aby se tomuto řešení vyhnuli – jednak proto, že jediné administrativní centrum činí síť zranitelnou, a pak také proto, že jediné centrum je limitujícím faktorem pro další růst. Bob Kahn, v té době jeden z ředitelů projektu, nesouhlas s takovým řešením vtipně komentoval: Centralizované řešení, mnohem jednodušší na řízení a kontrolování, byla populární mezi klasickými operátory telekomunikací. Ostatně jsou populární dodnes. „Chcete-li uspět při konstrukci sítě,“ říkával Bob Kahn, „je důležité sledovat, jak to dělá správa pošt a telekomunikací. Je dokonce důležité sledovat to velmi pozorně, abyste správně pochopili jejich rozhodnutí. A pak musíte udělat přesný opak.“ [...]
[...] Zjednodušeně je možno říci, že klasické telekomunikační firmy se snaží dodat služby co nejspolehlivější a co nejkvalitnější. [...] Snaží se dodat zaručené služby, aby tím mohli odůvodnit svoje nadsazené tarify, které by rády udrželi i nadále. Čili snaží se mít síť co nejvýkonnější, byť by se kvůli tomu měla stát velmi složitá. Na doporučení Boba Kahna Internet postupije přesně naopak. Důležité je zachovat síť co nejjednodušší a rozhodně do ní nikdy nepřidávat funkci, která by nakonec mohla být realizována přímo počítači-klienty. [...] Každý dodavatel Internetu je zároveň závislý i nezávislý na všech ostatních. Zájmem všech je co nejlépe rozvinout jejich síť a nybídnout levnější a výkonnější služby než jejich konkurenti. Všichni mají zájem se propojit a všichni to samozřejmě také dělají. Neexistuje komerční autorita, která by Inetrnetu vládla, ani centrální bod, odkud by se dalo kontrolovat veškerý provoz a přístupová práva. Ale naproti tomu mají všichni zájem spolupracovat, udržet globální propojení. Proto je nutné zaručit koherenci sítě, což vyžaduje přítomnost silné technické autority. Je nutné, aby nějaký centrální registr garantoval jednotu jmen a adres a aby nějaká centrální organizace stanovila normy pro přenášení dat na Internetu. [...]
[...] Ve Francii dnes máme telekomunikační monopol. Je pravda, že tento monopol se pokusil zamaskovat jako obyčejná firma tím, že se překřtil na France Telecom. Ale je to vážně podivná firma, když většina jejích zaměstnanců jsou úředníci. [...]Technické znalosti tu hrají jakous takous roli, ale ne rozhodující, protože z toho, kdo se příliš věnuje technice, se podle všeho nemůže stát dobrý ředitel. [...] Pokud chceme, aby francouzský telekomunikační průmysl přežil, musíme obchodovat s celým světem. Rovněž je třeba, aby telekomunikační poplatky byly na úrovni ostatních zemí, jestliže nechceme znevýhodnit ostatní sektory francouzského průmyslu. V současnosti k tomu máme stále velmi daleko, ačkoliv víme, že díky Evropské unii jsou to poslední léta telekomunikačních monopolů; ty by totiž po roce 1998 už neměly existovat. [...]
[...] Zrušení monopolu by mohlo uvolnit značné kapacity. V prvním okamžiku by měly podstatně klesnout ceny linek. [...] Při stejných investicích na jednoho uživatele kvalita sítě přímo závisí na počtu uživatelů. Vychází to ze samotného principu posílání paketů na Internetu: všichni uživatelé sdílejí všechny zdroje. ... spustí se známá spirála: růst sítě přinese zlepšení kvality a tím přitáhne další zájemce.
(Christian Huitema: A Bůh stvořil Internet... Mladá fronta, Praha 1996)
O autorovi
Oldřich Nečas
Doporučujeme

Dějiny psané čtyřmi písmeny 

Kde najdeme azbest
