










Fototrofní bakterie
Oceány pokrývají 70 % povrchu Země a ve vývoji života na naší planetě hrály vždy nezastupitelnou roli. Pochody probíhající v oceánech mají velký vliv na globální biologické i klimatické procesy. Ve světovém oceánu je přítomno velké množství bakterií, zhruba 3×1028, tj. řádově stomilionkrát více než hvězd ve viditelné části Vesmíru. Přitom o životě těchto bakterií toho víme velmi málo.
Nejdůležitější místo v mořském ekosystému zaujímají fotosyntetické mikroorganizmy, nazývané souborně fytoplankton. Využívají světelné záření jako základní zdroj energie, vážou oxid uhličitý do organických sloučenin a produkují kyslík (o fotosyntéze viz Vesmír 75, 33, 1996/1). Odhaduje se, že mořský fytoplankton vyrobí ročně asi 45 až 55 Pg (petagramů, 1 Pg je 1015 gramů) organického uhlíku, tedy přibližně stejné množství jako suchozemské rostliny (viz tabulku). Na této produkci organického uhlíku jsou závislé téměř všechny ostatní formy života v oceánu, heterotrofní bakterie, archea a eukaryonta. 1)
organizmů známe
Hlavní skupinou využívající zářivou energii jsou oxygenní fotosyntetické organizmy, tj. druhy produkující kyslík. Do této skupiny náležejí zelené rostliny, zelené či hnědé řasy, ruduchy a prokaryontní sinice. Přeměna světelné energie na energii chemickou zde probíhá souběžně ve dvou typech pigment-proteinových komplexů (ve fotosystému 1 a fotosystému 2). Světlo je nejdříve zachyceno ve světlosběrných komplexech obsahujících chlorofyl, a potom využito k rozdělení nábojů, které probíhá v reakčních centrech. Při osvětlení vytváří fotosyntetický aparát usměrněný tok elektronů. Ten slouží k syntéze NADPH + H+ (redukovaného nikotinamid adenin dinukleotid fosfátu), a následně je využit k asimilaci oxidu uhličitého. Zároveň s elektronovým transportem se na fotosyntetických membránách vytváří gradient protonů, který využívá enzymový komplex ATPáza. Výsledným produktem je ATP (adenozintrifosfát), který slouží jako „akumulátor“ energie pro biochemické reakce.
Druhou skupinou jsou anoxygenní fotosyntetické bakterie (nevyvíjející kyslík), které obsahují vždy pouze jeden typ reakčních center tvořených bakteriochlorofylem. Zdrojem elektronů v anoxygenní fotosyntéze jsou různé redukované látky, např. H2S, Fe2+, jednoduché organické látky aj. Podobně jako při oxygenní fotosyntéze slouží světelné reakce bakteriální fotosyntézy k produkci NADH + H+ a ATP. Anoxygenní fotosyntetické bakterie jsou vývojově velmi staré. Na Zemi se objevily přibližně před 3,5 miliardy let, tedy ještě před vznikem kyslíkové atmosféry, a většina z nich je proto na přítomnost kyslíku velmi citlivá. Po rozvoji oxygenních sinic a okysličení zhruba před 2 miliardami let ustoupily do specifických ekologických nik velmi omezených rozměrů. Dnes je nacházíme v sirných pramenech, ve vrstvách půdy s nedostatkem kyslíku, ve spodních bezkyslíkatých vrstvách sladkovodních jezer či ve spodní anaerobní vrstvě Černého moře.
Ještě je známa třetí skupina organizmů, které dokážou přímo využívat energii světelného záření. Jsou to fototrofní archea obsahující rodopsin. Na konci šedesátých let 20. století se zjistilo, že červené membrány extrémně slanomilných archeí rodu Halobacterium obsahují bakteriorodopsin (viz rámeček). Primární fotochemická reakce je u bakteriorodopsinu zcela jiného typu než fotosyntéza založená na (bakterio)chlorofylových reakčních centrech. Stejně jako fotosyntetické bakterie patří mezi vývojově velmi staré organizmy i fototrofní halobakterie. Dnes obývají již jen velmi specializované ekologické niky.
Vzhledem k značně omezenému rozšíření byl příspěvek fotosyntetických bakterií a fototrofních archeí ke globálním tokům energie a uhlíku považován za zanedbatelný. Výsledky práce dvou amerických oceánografických laboratoří však ukazují, že tyto představy bude zřejmě třeba přehodnotit.
Pracovníci laboratoře E. F. DeLonga v kalifornském Monterey Bay Aquarium Research Institute se rozhodli prověřit, zda se fototrofie, doposud známá jen u archeí, nevyskytuje i u mořských bakterií (eubakterií). Ve vzorcích DNA bakteriálního kmene SAR86, který byl izolován z Montereyského zálivu, skutečně nalezli sekvence podobné již dříve popsaným bakteriorodopsinovým genům. Fylogenetická analýza prokázala, že se nalezené sekvence od sekvencí popsaných pro bakteriorodopsin u archeí poněkud liší. Proto byl nový pigmentproteinový komplex pojmenován proteorodopsin. Zbývalo ovšem ještě prověřit, zda je nalezený gen skutečně schopný vytvořit funkční proteinový komplex. Nalezené geny byly proto přeneseny do heterotrofní bakterie E. coli, kde se z exprimovaných genů po přidání retinalu podařilo sestavit funkční fotochemické komplexy. Po osvětlení začaly proteorodopsinové komplexy přečerpávat protony přes membránu. První krok k přeměně světelné energie v energii chemickou se uskutečnil.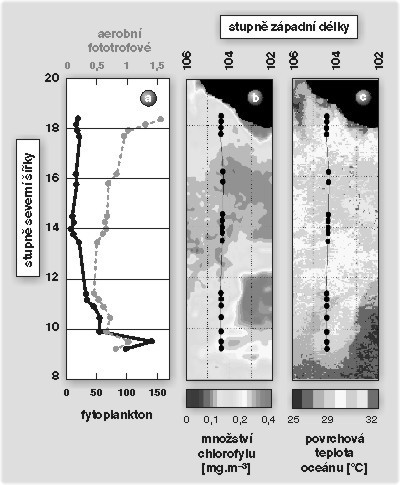
Po získání těchto laboratorních důkazů se tým zaměřil na vzorky přirozených bakteriálních společenstev odebraných v Montereyském zálivu. Osvětlení vzorků intenzivním laserovým zábleskem vyvolalo v absorpčním spektru obdobnou změnu, jakou projevovaly komplexy proteorodopsinu v buňkách E. coli.
Další výzkum prokázal přítomnost proteorodopsinových genů i u jiných bakteriálních společenstev, která byla odebrána v různých částech Tichého oceánu, na Havaji a v Antarktidě. To napovídá, že proteorodopsinová fotosyntéza je v oceánech široce rozšířena.
Ani heterotrofie, ani fototrofie – fotoheterotrofie
Na opačné straně Spojených států, na Rutgersově univerzitě v New Jersey, se tým P. Falkowského již mnoho let zabývá vývojem optických metod měření fotosyntézy v oceánu. Vyvinul řadu přístrojů pro měření fluorescenční emise chlorofylu, která slouží jako specifický ukazatel přítomnosti a funkce fotosyntetických organizmů. Před třemi lety zaujala P. Falkowského hypotéza, podle níž se předpokládá vznik nejprimitivnějších fotosyntetických organizmů v blízkosti hlubokomořských sopečných vývěrů. Specifické chemické prostředí spolu s tepelným zářením uvolňovaným z vývěrů zde mohlo vytvořit vhodné podmínky pro vznik primitivní formy fotosyntézy, založené na bakteriochlorofylu. Na rozdíl od chlorofylů totiž bakteriochlorofyly dokážou využívat také infračervené záření o vlnových délkách 750 až 1100 nm. Po rychlém vývoji přístroje schopného zaznamenat až pikomolární koncentrace bakteriochlorofylu se tým vypravil do tektonicky aktivních míst východního Tichého oceánu. Pro měření byl využit hlubokomořský batyskaf Alvin z oceánografického ústavu ve Woods Hole. Ani po několikátém sestupu do hloubky dvou kilometrů k sopečným vývěrům však fluorimetr přítomnost fotosyntetických organizmů nezaregistroval. Znejistěný tým, obávající se závady, přezkoušel činnost přístroje s využitím mořské vody odebrané z povrchových vrstev moře nad místem ponoru Alvinu. Předpokládalo se, že v povrchových vodách bude přítomen jen fytoplankton emitující ve spektrální oblasti chlorofylu, zatímco ve spektrální oblasti bakteriochlorofylu nebude emise žádná. Přístroj ale zaznamenal silný signál bakteriochlorofylu (indikující přítomnost fotosyntetických bakterií) právě ve vzorku povrchové vody. Při dalších měřeních se zjistilo, že se signál bakteriochlorofylu vyskytuje v povrchové prosvětlené vrstvě oceánu do hloubky zhruba 100 m a je možné ho sledovat po celé délce dvanáctisetkilometrového transektu Tichým oceánem, který byl mapován v průběhu plavby (obr. 2). Poměr množství bakteriochlorofylu k množství chlorofylu se ve sledované oblasti pohyboval mezi 1 až 5 %.Při dalších plavbách se v severovýchodním Tichomoří podařilo kvantifikovat podíl fotosyntetických bakterií v eutrofní vrstvě pomocí fluorescenční mikroskopie. V těchto oblastech chudých na živiny představovaly fotosyntetické bakterie až 10 % celkové bakteriální komunity.
Později se z odebraných vzorků vody podařilo izolovat v laboratoři několik kmenů aerobních fotosyntetických bakterií rodu Erythrobacter. Aerobní fotosyntetické bakterie jsou fylogeneticky příbuzné anaerobním purpurovým bakteriím, s nimiž sdílejí stejné základní fotosyntetické pigmenty i architekturu reakčních center. Na rozdíl od nich se však v průběhu evoluce přizpůsobily kyslíkové atmosféře a staly se výhradními aeroby. Podobně jako běžné heterotrofní bakterie vyžadují i tyto zdroj organického uhlíku pro růst, navíc však dokážou – omezeně – vázat oxid uhličitý. Fotosyntéza jim pravděpodobně slouží jako významný doplňkový zdroj energie, který je schopen pokrýt až 50 % energetických potřeb buňky. Podle odhadů mohou aerobní fotosyntetické bakterie přispívat zhruba 1 až 3 % do úhrnného množství zářivé energie přeměněné mořskými fotosyntetickými organizmy na energii metabolickou.
Tato nová zjištění rozšiřují naše představy o metabolických strategiích a energetických tocích v mořské bakteriální komunitě. Opravují dřívější jednoduché schéma, které dělí mořské organizmy na fytoplankton, tj. primární fotosyntetické producenty organického uhlíku, a heterotrofní organizmy, jež organický uhlík využívají. Zmiňované typy fototrofních bakterií zaujímají pozici mezi oběma hlavními skupinami (obr. 3). Podobně jako fytoplankton využívají energii získanou ze světla, podobně jako heterotrofové jsou však závislé na zdrojích organického uhlíku. Tato možnost pokrýt část metabolických požadavků zachycováním světelné energie je v oceánských oblastech s velmi omezenými zdroji živin výhodnou strategií. 3)
Literatura
Béja O., Spudich E. N., Spudich J. L., Leclerc M., DeLong E. F.: Proteorhodopsin phototrophy in the ocean, Nature 411, 786–789, 2001Copley J.: All at sea, Nature 415, 572–574, 2002
Kolber Z. S., Plumley F. G., Lang A. S., Beatty J. T., Blankenship R. E., VanDover C. L., Vetriani C., Koblížek M., Rathgeber C., Falkowski P. G.: Contribution of aerobic photoheterotrophic bacteria to the carbon cycle in the ocean, Science 292, 2492–2495, 2001
Poznámky
Bakteriorodopsin je fotochemický komplex slanomilných archeí rodu Halobacterium. Skládá se z pigmentu retinalu, navázaného na membránový protein (opsin). Bakteriorodopsin funguje jako jednoduché čerpadlo, které v cyklické fotochemické reakci přenáší vodíkové ionty přes membránu. Vytvořený gradient poté pohání enzymový komplex ATPázy, produkující ATP (adenozintrifosfát). Strukturní analog bakteriorodopsinu – rodopsin – je dobře znám u živočichů, kde představuje základní světločivý prvek v oční sítnici.
Bakteriochlorofyl je tetrapyrolové barvivo nacházející se ve fotosyntetických bakteriích. Je známo několik forem bakteriochlorofylu (označují se písmeny a až g), které se mírně liší v chemické struktuře a spektrálních vlastnostech. Po zachycení světelné energie dochází v reakčních centrech k rozdělení nábojů. Redukční potenciál vzniklých elektronů je v metabolizmu využit pro fixaci oxidu uhličitého. Zároveň se přes membránu přenášejí vodíkové ionty, které – stejně jako u bakteriorodopsinu – slouží k produkci ATP.
O autorech
Michal Koblížek
Ondřej Prášil
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
