Teorie ideálního lesa
| 9. 6. 2011 Chceme-li porozumět něčemu složitému, máme řadu možností, jak k tomu přistoupit. Můžeme začít popisovat jednotlivé složky, třídit je a snažit se vystihnout vztahy mezi nimi. Tak to většinou dělá biologie. Fyzika postupuje jinak. Snaží se postihnout základní rysy studované skutečnosti pomocí idealizovaného systému (jako je třeba hmotný bod, ideální plyn) a pozorovanou rozmanitost vysvětlovat jako nejrůznější odchylky od ideální představy skutečnosti. Tento postup se velmi osvědčil zatím hlavně právě ve fyzice. Začínají se však objevovat pokusy použít podobný postup ve vědách o živé přírodě.
Chceme-li porozumět něčemu složitému, máme řadu možností, jak k tomu přistoupit. Můžeme začít popisovat jednotlivé složky, třídit je a snažit se vystihnout vztahy mezi nimi. Tak to většinou dělá biologie. Fyzika postupuje jinak. Snaží se postihnout základní rysy studované skutečnosti pomocí idealizovaného systému (jako je třeba hmotný bod, ideální plyn) a pozorovanou rozmanitost vysvětlovat jako nejrůznější odchylky od ideální představy skutečnosti. Tento postup se velmi osvědčil zatím hlavně právě ve fyzice. Začínají se však objevovat pokusy použít podobný postup ve vědách o živé přírodě.
Ta je ovšem na první pohled mnohem složitější než fyzikální systémy, proto je třeba vybrat si pro začátek nějaký jednodušší systém. Paradoxně ale právě systém, který je považován za archetyp složitosti, totiž tropický deštný prales, posloužil k vybudování teorie založené na jednoduchých principech fungování idealizovaného systému. Složitost je totiž věcí nazírání, nikoli přírody samotné. Příroda je složitá vždycky, jde jen o to, co si vybereme, kterým fenoménům chceme porozumět a co zanedbáme nebo budeme považovat za šum.
 Existuje řada důvodů, proč začít zrovna pralesem. Les je vlastně ultimátním suchozemským prostředím, které nakonec převládne na každém místě, kde nepůsobí omezující faktory, jako je zima, sucho nebo nedostatečně hluboký substrát (obr. 1). Je tomu tak proto, že při vývoji suchozemské vegetace vítězí rostliny, které přerostou a zastíní ty ostatní – a to dokážou nejlíp právě stromy. Jejich trik spočívá v tom, že podpůrná struktura, tedy kmen, díky němuž se dostanou výš než ostatní, je tvořena z větší části mrtvým pletivem, do jehož udržení nemusejí dále investovat energii. Naproti tomu větší byliny musejí investovat do stonku z živých buněk tolik energie, že už to nakonec „neutáhnou“. Dřeviny, a zvláště stromy tak vítězí, kdykoli jim to podmínky umožňují. Vývoj vegetace přirozeně spěje k lesu, jen k němu někdy nedospěje. Proto je dobré začít právě lesem jakožto prostředím, jehož přítomnost nemusíme vysvětlovat nějakými specifickými podmínkami.
Existuje řada důvodů, proč začít zrovna pralesem. Les je vlastně ultimátním suchozemským prostředím, které nakonec převládne na každém místě, kde nepůsobí omezující faktory, jako je zima, sucho nebo nedostatečně hluboký substrát (obr. 1). Je tomu tak proto, že při vývoji suchozemské vegetace vítězí rostliny, které přerostou a zastíní ty ostatní – a to dokážou nejlíp právě stromy. Jejich trik spočívá v tom, že podpůrná struktura, tedy kmen, díky němuž se dostanou výš než ostatní, je tvořena z větší části mrtvým pletivem, do jehož udržení nemusejí dále investovat energii. Naproti tomu větší byliny musejí investovat do stonku z živých buněk tolik energie, že už to nakonec „neutáhnou“. Dřeviny, a zvláště stromy tak vítězí, kdykoli jim to podmínky umožňují. Vývoj vegetace přirozeně spěje k lesu, jen k němu někdy nedospěje. Proto je dobré začít právě lesem jakožto prostředím, jehož přítomnost nemusíme vysvětlovat nějakými specifickými podmínkami.
Věková a velikostní struktura pralesa
 Ocitneme-li se v opravdovém pralese, můžeme si všimnout nápadné věci, která jako by byla v rozporu s tradiční představou lesa složeného z prastarých obřích velikánů. Ne že by tu takové stromy nebyly, ale rozhodně nepřevažují – převažují naopak útlé kmínky, a čím jsou útlejší, tím více převažují (obr. 2). Když se nad tím zamyslíme, není čemu se divit. Pokud je les stabilní, musí se neustále obnovovat; musí tam být tedy mladší jedinci, kteří časem nahradí ty starší. A vzhledem k tomu, že během růstu někteří jedinci umírají, mladších jedinců musí být víc než těch starších.
Ocitneme-li se v opravdovém pralese, můžeme si všimnout nápadné věci, která jako by byla v rozporu s tradiční představou lesa složeného z prastarých obřích velikánů. Ne že by tu takové stromy nebyly, ale rozhodně nepřevažují – převažují naopak útlé kmínky, a čím jsou útlejší, tím více převažují (obr. 2). Když se nad tím zamyslíme, není čemu se divit. Pokud je les stabilní, musí se neustále obnovovat; musí tam být tedy mladší jedinci, kteří časem nahradí ty starší. A vzhledem k tomu, že během růstu někteří jedinci umírají, mladších jedinců musí být víc než těch starších.
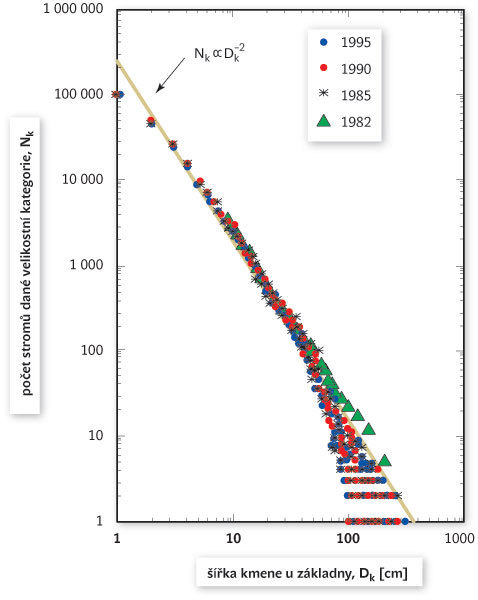
Tak tomu v rovnovážných přirozených pralesích skutečně je. A nejen to. Když se podíváme na frekvenční rozložení různých velikostních tříd stromů, najdeme pozoruhodnou zákonitost. Vyneseme-li na vodorovnou osu šířku stromu v určité (malé) výšce nad zemí (která za předpokladu rovnoměrného přírůstku stromu do šířky za jednotku času je zároveň úměrná věku stromu) a na svislou osu počet stromů na jednotku plochy v dané velikostní kategorii, obojí v logaritmickém měřítku, objeví se nám přímka, která má směrnici –2 (obr. 3). Tohle platí nejen pro tropické, ale i pro nejrůznější jiné přirozené lesy, tedy třeba také pro pralesy mírného pásu.1) Má to jeden zajímavý důsledek. Kdybychom na vodorovnou osu vynesli namísto šířky průřez (tedy plochu potenciálního řezu), dostaneme také přímku v logaritmicko-logaritmickém měřítku, tentokrát ale se sklonem –1, poněvadž plocha kruhu je úměrná druhé mocnině průměru (viz rámeček). Tento vztah je vlastně obyčejná nepřímá úměra: oč mají stromy menší průřez, o to jsou početnější. Z toho plyne, že když sečteme všechny průřezy stromů dané velikostní kategorie v dané oblasti, vyjde nám vždycky to samé číslo: celkový průřez stromů dané věkové kategorie je nezávislý na věku.
 Právě toto pozorování vedlo autory metabolické teorie biologie (viz Vesmír 83, 508, 2004/9 a Vesmír 89, 536, 2010/9) k vybudování obecné teorie struktury a dynamiky přirozeného lesa.2) Je vcelku jednoduchá. Předpokládá jen určitou pravidelnost v dávno známém jevu zvaném samozřeďování. Jak stromky rostou, postupně některé odumírají, takže do další věkové kategorie se jich dostane jen část, jejíž celkový průřez všech kmenů je stejný jako u předchozího stadia. Souvisí to s tím, že čím větší je průřez kmene, tím větší je tok vody a živin cévními systémy, a tím je tedy intenzivnější celkový metabolismus stromu. Průřez kmene je tak úměrný metabolismu a konstantní suma průřezů dané věkové kategorie tedy zároveň znamená, že každá věková kategorie spotřebuje přibližně stejné množství energie. Podobný jev (který nazýváme energetická ekvivalence) pozorujeme i u vysázených monokultur: celková spotřeba energie na jednotce plochy nezávisí na velikosti jednotlivých rostlin, poněvadž o co jsou jedinci větší, o to je jich míň (obr. 4).
Právě toto pozorování vedlo autory metabolické teorie biologie (viz Vesmír 83, 508, 2004/9 a Vesmír 89, 536, 2010/9) k vybudování obecné teorie struktury a dynamiky přirozeného lesa.2) Je vcelku jednoduchá. Předpokládá jen určitou pravidelnost v dávno známém jevu zvaném samozřeďování. Jak stromky rostou, postupně některé odumírají, takže do další věkové kategorie se jich dostane jen část, jejíž celkový průřez všech kmenů je stejný jako u předchozího stadia. Souvisí to s tím, že čím větší je průřez kmene, tím větší je tok vody a živin cévními systémy, a tím je tedy intenzivnější celkový metabolismus stromu. Průřez kmene je tak úměrný metabolismu a konstantní suma průřezů dané věkové kategorie tedy zároveň znamená, že každá věková kategorie spotřebuje přibližně stejné množství energie. Podobný jev (který nazýváme energetická ekvivalence) pozorujeme i u vysázených monokultur: celková spotřeba energie na jednotce plochy nezávisí na velikosti jednotlivých rostlin, poněvadž o co jsou jedinci větší, o to je jich míň (obr. 4).
Další podivnosti
 Skutečnost, že všechny věkové kategorie stromů v pralese spotřebovávají přibližně stejně energie, má další zajímavé konsekvence. Na rozdíl od vžité představy pralesa charakterizovaného hustým korunovým patrem, pod nímž je volno a tma, bývá celý prostor celkem rovnoměrně vyplněn listím, poněvadž i malé stromky musí fotosyntetizovat, a malých je tam spousta – o to víc, oč jsou menší. Celková listová plocha je úměrná metabolismu, a tedy také oné sumě průřezů, čili rovněž nezávisí na velikosti stromů. Místo abychom pozorovali jednolitý a rovnoměrný baldachýn v určité výšce (nebo několik dobře odlišitelných pater v určitých výškách), vidíme spíše stromy nejrůznějších velikostí, které netvoří nijak pěkně oddělená patra, mezery jsou občas vyplněné korunami menších stromů, zkrátka nepořádek (obr. 5). V našich podmínkách se s něčím podobným setkáme pouze v takzvaném stadiu rozpadu pralesa. V nížinném tropickém pralese jako by byl rozpad všudypřítomný. Vskutku se způsob, jak se prales obnovuje, podobá trvalému rozpadu.
Skutečnost, že všechny věkové kategorie stromů v pralese spotřebovávají přibližně stejně energie, má další zajímavé konsekvence. Na rozdíl od vžité představy pralesa charakterizovaného hustým korunovým patrem, pod nímž je volno a tma, bývá celý prostor celkem rovnoměrně vyplněn listím, poněvadž i malé stromky musí fotosyntetizovat, a malých je tam spousta – o to víc, oč jsou menší. Celková listová plocha je úměrná metabolismu, a tedy také oné sumě průřezů, čili rovněž nezávisí na velikosti stromů. Místo abychom pozorovali jednolitý a rovnoměrný baldachýn v určité výšce (nebo několik dobře odlišitelných pater v určitých výškách), vidíme spíše stromy nejrůznějších velikostí, které netvoří nijak pěkně oddělená patra, mezery jsou občas vyplněné korunami menších stromů, zkrátka nepořádek (obr. 5). V našich podmínkách se s něčím podobným setkáme pouze v takzvaném stadiu rozpadu pralesa. V nížinném tropickém pralese jako by byl rozpad všudypřítomný. Vskutku se způsob, jak se prales obnovuje, podobá trvalému rozpadu.
 Budiž ovšem řečeno, že takovéto rozvrstvení- nerozvrstvení, kdy všechny velikostní kategorie jsou stejně zastoupené a listí je rovnoměrně rozmístěno všude, už není typické například pro horské tropické pralesy, které jsou charakteristické spíš obřími stromy s rozsochatými větvemi obrostlými celými zahradami epifytů (obr. 6), pod nimiž je spousta prostoru a až u země najdeme hustý podrost s množstvím kapradin, včetně stromových. To ale lze interpretovat tak, že horský prales už není tím ideálním lesem a uplatňují se v něm nějaké jiné procesy. Tak je tomu i u řady dalších lesů, jak si ještě povíme.
Budiž ovšem řečeno, že takovéto rozvrstvení- nerozvrstvení, kdy všechny velikostní kategorie jsou stejně zastoupené a listí je rovnoměrně rozmístěno všude, už není typické například pro horské tropické pralesy, které jsou charakteristické spíš obřími stromy s rozsochatými větvemi obrostlými celými zahradami epifytů (obr. 6), pod nimiž je spousta prostoru a až u země najdeme hustý podrost s množstvím kapradin, včetně stromových. To ale lze interpretovat tak, že horský prales už není tím ideálním lesem a uplatňují se v něm nějaké jiné procesy. Tak je tomu i u řady dalších lesů, jak si ještě povíme.
 Z ideálního lesa, charakterizovaného zmíněnými symetriemi, můžeme odvodit některé další skutečnosti. Ze zákonitosti týkající se hustoty stromů dané velikosti lze odvodit, že průměrná vzdálenost mezi stromy dané velikosti bude úměrná šířce stromu (viz rámeček): když budeme tedy vědět, že dejme tomuprůměrná vzdálenost k nejbližšímu sousedu stejné velikosti bude u stromu s deseticentimetrovou šířkou kmene pět metrů, můžeme předpokládat, že mezi nejbližšími dvaceticentimetrovými kmeny bude v průměru vzdálenost deset metrů. Známe-li navíc vztahy mezi tloušťkou kmene a rozměrem koruny, můžeme odvodit, že větší stromy budou mít koruny vzájemně izolovanější – a odtud právě pramení onen zmíněný pocit „nepořádku“ v pralese.
Z ideálního lesa, charakterizovaného zmíněnými symetriemi, můžeme odvodit některé další skutečnosti. Ze zákonitosti týkající se hustoty stromů dané velikosti lze odvodit, že průměrná vzdálenost mezi stromy dané velikosti bude úměrná šířce stromu (viz rámeček): když budeme tedy vědět, že dejme tomuprůměrná vzdálenost k nejbližšímu sousedu stejné velikosti bude u stromu s deseticentimetrovou šířkou kmene pět metrů, můžeme předpokládat, že mezi nejbližšími dvaceticentimetrovými kmeny bude v průměru vzdálenost deset metrů. Známe-li navíc vztahy mezi tloušťkou kmene a rozměrem koruny, můžeme odvodit, že větší stromy budou mít koruny vzájemně izolovanější – a odtud právě pramení onen zmíněný pocit „nepořádku“ v pralese.
Nejpodivnější ale je, že tytéž zákonitosti, které se týkají jednotlivých různě velkých kmenů a kmínků, lze uplatnit i na jednotlivé větve v rámci stromu jediného i na větve v rámci celého lesa. Větve se postupně dělí do tenčích větviček, kterých je přesně podle zmíněného pravidla o to víc. Tenčí větve jsou tak ekvivalentní tenčím kmínkům; liší se od nich jen tím, že nerostou rovnou ze země. Když se nad tím zamyslíme, není to až tak překvapivé: celkový průřez vodivých pletiv, kterému odpovídá tok látek cévním systémem stromu, zůstává v rámci stromu víceméně zachován. Tohle pravidlo mimochodem objevil už Leonardo da Vinci a zdá se, že opravdu platí pro většinu rostlin. Všude ovšem najdeme nějaké výjimky.
Suchozemská příroda jako poruchy ideálního lesa
Čtenář, který o lese něco ví, může teď začít protestovat, že většina lesů takhle jednoduše nefunguje – a má pravdu. Mluvili jsme skutečně o ideálním lese, jemuž se sice řada přirozených pralesů nějak blíží, ale v čisté podobě ho sotva někde najdeme. Přesto nám tato idealizovaná představa (která původně nebyla vytvořena z čistě teoretického modelu, ale spíše abstrakcí z pravidelností pozorovaných v reálných pralesích) může být vodítkem pro to, co lze považovat za „správný les“. Ukázalo se například, že různé lesy se odchylují od oné univerzální zákonitosti (obr. 3) podle toho, jak daleko jsou od klimaxového, tedy vyspělého rovnovážného stadia. Na začátku ekologické sukcese, tedy přirozeného vývoje daného prostředí, je pro porosty typičtější mnohem strmější sklon závislosti mezi šířkou kmene a početností, jinými slovy je tam daleko větší převaha užších kmínků (což je logické, když jde o mladý les). Teprve když les vyspěje, ustaví se rovnováha charakterizovaná sklonem –2. Takový les svou strukturu už dál nemění.
Čím více se daný porost odlišuje od ideálního lesa, tím více dalších faktorů potřebujeme, abychom vysvětlili jeho strukturu. Potkáme-li vysokokmenný majestátní porost vzrostlých stromů, třeba vzrostlou bučinu (které se občas celkem případně říká chrámová bučina), můžeme předpokládat, že tu asi není moc dlouho – musela vzniknout jednorázovým vysazením či stejnověkým náletem – a že asi v tomhle stavu dlouho nevydrží, neboť tu nejsou jedinci, kteří by vzrostlé dospělé stromy časem nahradili. Odchylky od ideální struktury ale nemusí nutně indikovat lidský management; podobnou strukturu mají i přirozené lesy s požárovou dynamikou, jako je tajga. Les pak prostě roste, dokud se tam nenahromadí tolik dřevní hmoty, že rázem shoří, a vše začíná nanovo skoro od nuly. Jinými slovy, ani zásadní odchylky od ideálního lesa neznamenají, že nejde o přirozený a „správný“ les; jen musíme zjistit, co odchylku způsobilo.
Když to trochu zobecníme, veškerou suchozemskou vegetaci můžeme považovat za různé poruchy ideálního lesa. Když budou podmínky ideální, vyroste postupně les charakterizovaný zmíněnými zákonitostmi. Povede-li sezonalita prostředí k tomu, že v některých obdobích bude v lese málo živé, biologicky aktivní hmoty, bude mít les větší tendenci opakovaně hořet, takže se budou střídat stejnověká stadia. Když se k tomu přidá sucho, bude hořet tak často, že ani žádný pořádný les nevyroste, maximálně roztroušené křoví a ojedinělé stromy; jakmile by se porost zapojil, pravděpodobnost požáru by vzrostla a porost by se zas rozpojil. Tak funguje třeba africká savana s výjimkou těch nejsušších oblastí: nízký les by potenciálně mohl růst skoro všude, ale požáry mu to nedovolí. V ještě sušších oblastech už ale nedostatek vody natolik omezí růst stromů, že převládnou jiné růstové formy. K tomu mohou přispět i býložravci, jejichž okus je mnohem fatálnější pro dřeviny než třeba pro trávy. Sami býložravci však nejsou přirozeně schopni zlikvidovat pořádný vlhký a druhově bohatý les, pokud jim nepomohou další faktory.
Diverzita lesa
Zatím jsme uvažovali jen o věkové struktuře lesa a nepřemýšleli, jak je to s různými druhy. Platí v rámci každého druhu stejné zákonitosti jako pro celý les? To asi sotva; víme, že jsou dřeviny charakterizující jen nižší stromové patro a že jen některé druhy vyrostou do plné výše. Druhové složení malých kmínků může být tedy odlišné od složení velkých kmenů. Jak funguje dynamika populací jednotlivých druhů, aby dohromady složily ono pozorované velikostní rozložení a aby zároveň jeden druh časem nepřerostl všechny ostatní, není dosud pořádně prozkoumané. Mechanismy udržování druhové rozmanitosti lesa v jednom typu prostředí (když si nevystačíme s tím, že každý druh prospívá lépe na jiném typu substrátu) známe kupodivu o hodně lépe z tropických pralesů než z našich listnatých lesů, poněvadž tento problém představoval centrální záhadu tropické biologie. Tropičtí biologové řešili po mnoho desetiletí, jak je možné, že na pár hektarech mohou růst až stovky druhů stromů a většina z nich jen v jednom nebo dvou exemplářích; jak je možné, že jeden nebo několik málo druhů nevytlačí všechny ostatní? Dnes se zdá, že hlavním mechanismem udržování diverzity je frekvenčně závislé přežívání, spočívající v tom, že semenáčky a obecně mladší jedinci špatně přežívají v přítomnosti starších jedinců téhož druhu. Jakmile se tedy jeden druh trochu namnoží, začnou být značně znevýhodněni jeho potomci – a naopak, vyplatí se být vzácný, poněvadž potomci vzácného stromu budou mít výrazně vyšší šanci dožít se dospělosti. Tenhle jednoduchý mechanismus zaručí udržování velmi vysoké druhové rozmanitosti.
Nikdo zatím netuší, co tento efekt zvýhodňující vzácnost způsobuje – snad druhově specifické parazitické houby, které se namnoží vždycky, když jeden druh začne převažovat, a napadají pak jeho potomky. Nikdo je ovšem zatím nenašel a svádíme to na ně zkrátka proto, že o houbách skoro nic nevíme. Každopádně má tento efekt zajímavé důsledky pro dynamiku pralesa. Vede například k tomu, že starší věkové kategorie jsou druhově vyrovnanější než ty mladší, poněvadž v těch mladších se ještě najdou potomci hojných druhů, kteří časem s větší pravděpodobností zemřou. Celkové početnosti jednotlivých druhů sice nakonec stejně nebudou vyrovnané (vždy převažuje jen pár druhů a ostatní jsou vzácné, viz Vesmír 88, 784, 2009/12), lze je ale předpovědět na základě toho, jak silně reagují semenáčky na přítomnost dospělců téhož druhu. Nejpočetnější druhy tropického lesa jsou totiž ty, jejichž potomkům nejmíň záleží na tom, kolik je kolem nich vzrostlých stromů téhož druhu.3)
Můžeme si tedy představit přirozený, druhově bohatý les jako časoprostorovou mozaiku, v níž se přelévají vlny představující jednotlivé druhy, přičemž každá vlna je následovaná dočasným ústupem daného druhu. Některé druhy přitom přežívají v malých počtech, jiné jsou četnější, ale jejich počty v daném místě oscilují a nikdy úplně nepřevládnou. Na každém místě to přitom bude trochu jiné v závislosti na tom, který druh se tam dočasně vzchopil. To vše ale probíhá při zachování celkové struktury pralesa, takže nezasvěcený pozorovatel ani nezaznamená, jak dramatické děje se tu odehrávají.
Poučení
Každý les je jiný a v každém jistě panují trochu jiné zákonitosti, ať už tím myslíme cokoli. Přesto existuje jakýsi úběžník, ideál, k němuž se lesy na různých místech zemského povrchu více či méně blíží. Ten se vyznačuje zajímavými symetriemi – a odchylky od těchto symetrií lze interpretovat jako následky nejrůznějších dalších vlivů. Pozoruhodné je, že nejvíc se k ideálnímu lesu blíží tropické deštné pralesy, které jsou v řadě ohledů skutečně nejsložitějším prostředím na Zemi. Bude to asi proto, že právě u složitého a zároveň stabilního systému se nejrůznější individuální vlivy (vlastnosti jednotlivých druhů, historie jednotlivých složek a povaha individuálních interakcí) vyruší a strukturu i dynamiku budou ovlivňovat především obecné rysy lesa. Složitost tak může být mnohem pevnějším základem budování obecných teorií než jednoduchost, která se projevuje vždy jinými konkrétními podmínkami a specifickým vývojem.
V poslední době řada autorů dokládá, že i ty nejopravdovější pralesy se řídí trochu jinými zákonitostmi; uvažují přitom o dostupnosti zdrojů, která se mění v závislosti na velikosti stromů a zpětné vazbě mezi stromy a prostředím.4) Jenže podobné zajímavosti lze odhalovat teprve tehdy, když máme teorii, která říká, jak by to mělo fungovat v tom nejjednodušším případě. Mít teorii ideálního lesa je dobré už jen proto, abychom věděli, vůči čemu se eventuálně vymezovat. Platí tak starý bonmot, že máloco je tak praktického jako dobrá teorie.
Poznámky
1) Enquist B. J. & Niklas K. J.: Invariant scaling relations across tree-dominated communities. Nature 410, 655–660, 2001.
2) West G. B., Enquist B. J., Brown J. H.: A general quantitative theory of forest structure and dynamics. PNAS 106: 7040–7045, 2009; Enquist B. J., West G. B., Brown J. H.: Extensions and evaluations of a general quantitative theory of forest structure and dynamics. PNAS 106, 7046–7051, 2009.
3) Comita L. S., Muller-Landau H. C., Aguilar S. & Hubbell S. P.: Asymmetric density dependence shapes species abundances in a tropical tree community. Science 329, 330–332, 2010.
4) Muller-Landau H. C. et al.: Testing metabolic ecology theory for allometric scaling of tree size, growth and mortality in tropical forest. Ecology Letters 9, 575–588, 2006; Muller-Landau H. C. et al.: Comparing tropical forest tree size distributions with the predictions of metabolic ecology and equilibrium models. Ecology Letters 9, 589–602, 2006.
Skutečnost, že rozložení četnosti různých velikostních tříd stromů v ideálním lese lze vyjádřit jako přímku v logaritmicko-logaritmickém měřítku (obr. 3) se sklonem –2, znamená, že vztah mezi četností a šířkou kmene D můžeme vyjádřit rovnicí přímky y = ax + b, tedy v tomto případě
logNk = logN0 – 2logDk,
kde Nk je počet stromů (na jednotku plochy) v dané velikostní třídě, N0 je počet těch nejtenčích stromů (respektive semenáčků, podle toho, u jaké velikosti začínáme) a –2 je sklon té přímky. Po odlogaritmování dostaneme
Nk = N0Dk–2.
Jde tedy o mocninné rozložení (viz též Vesmír 80, 490, 2001/9). Víme-li, že průřez kmenem P je úměrný druhé mocnině šířky P ~ D2, a tedy D~P½, můžeme odvodit, že Nk ~ (Pk½)–2, takže
Nk ~ Pk–1.
Počet jedinců na jednotku plochy je tedy nepřímo úměrný průřezu jejich kmenů. Chceme-li pak znát vztah mezi tloušťkou kmene a vzdáleností nejbližšího souseda o stejně tloušťce, stačí vědět, že průměrná vzdálenost L mezi jedinci je nepřímo úměrná odmocnině z hustoty jedinců, tedy Lk ~ Nk–½ (zdvojnásobení vzdáleností mezi jedinci vede k čtyřikrát menším hustotám).
Tím pádem
Lk ~ (Dk–2)–½ = Dk,
čili průměrná vzdálenost mezi jedinci dané velikosti je úměrná šířce kmenů.
Ke stažení
 článek ve formátu pdf [1,25 MB]
článek ve formátu pdf [1,25 MB]
O autorovi
David Storch

Doporučujeme

Rostliny vyprávějí o lidech

O prázdnech v nás