Přestavba buněčného jádra v tyčinkách sítnice:
Na úsvitu své evoluce prošli savci dlouhým obdobím noční aktivity. Adaptace na noční život výrazně ovlivnila jejich senzorickou výbavu. Na jedné straně došlo k velké diverzifikaci čichových receptorů a k mnohačetným zdokonalením sluchového aparátu, na straně druhé k výraznému omezení barevného vidění.
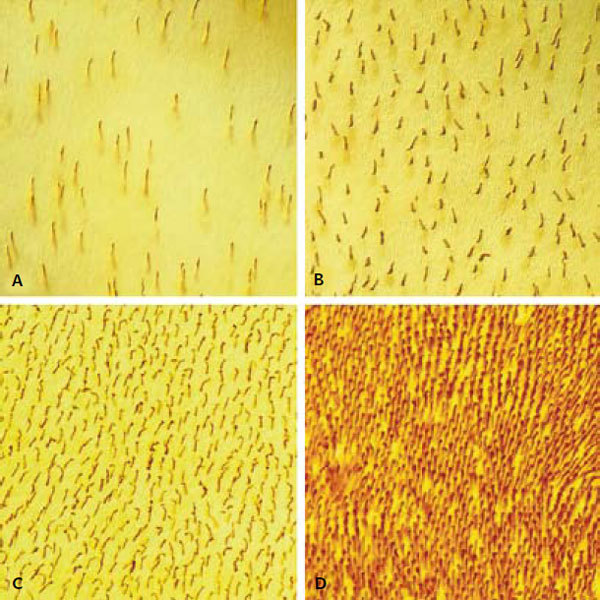 Většina obratlovců je vybavena tyčinkami a čtyřmi typy čípků, které jsou charakterizovány čtyřmi spektrálně odlišnými opsiny (světločivnými proteiny). Dva typy opsinů byly v rané evoluci savců nenávratně ztraceny. Většina savců má tedy pouze tři typy fotoreceptorů: tyčinky, které exprimují rodopsin a zabezpečují vidění při nízkých intenzitách světla (k „podráždění“ tyčinky dostačuje dopad několika fotonů), a dva typy čípků, které exprimují dva spektrálně různé opsiny a zabezpečují barevné vidění. Většina savců má tedy dichromatické barevné vidění, tj. jsou schopni rozeznat modrou barvu od zelené, ale nikoli zelenou od červené. Noční savci naopak vynikají schopností vidět za intenzit světla, které jsou milionkrát nižší než intenzity běžné během dne. Této citlivosti je dosaženo zvýšením hustoty tyčinek a snížením poměru čípků a tyčinek (obr. 1).1) Důsledkem zvýšení hustoty tyčinek je však enormní zvýšení počtu jader fotoreceptorů ve vnější jaderné vrstvě (obr. 2A), což vede ke zhoršení optických vlastností sítnice. Nedávno demonstrovali Irina Solovei a spol.2) v elegantní studii, že u nočních savců je tato nevýhoda kompenzována unikátní přestavbou jader tyčinek.
Většina obratlovců je vybavena tyčinkami a čtyřmi typy čípků, které jsou charakterizovány čtyřmi spektrálně odlišnými opsiny (světločivnými proteiny). Dva typy opsinů byly v rané evoluci savců nenávratně ztraceny. Většina savců má tedy pouze tři typy fotoreceptorů: tyčinky, které exprimují rodopsin a zabezpečují vidění při nízkých intenzitách světla (k „podráždění“ tyčinky dostačuje dopad několika fotonů), a dva typy čípků, které exprimují dva spektrálně různé opsiny a zabezpečují barevné vidění. Většina savců má tedy dichromatické barevné vidění, tj. jsou schopni rozeznat modrou barvu od zelené, ale nikoli zelenou od červené. Noční savci naopak vynikají schopností vidět za intenzit světla, které jsou milionkrát nižší než intenzity běžné během dne. Této citlivosti je dosaženo zvýšením hustoty tyčinek a snížením poměru čípků a tyčinek (obr. 1).1) Důsledkem zvýšení hustoty tyčinek je však enormní zvýšení počtu jader fotoreceptorů ve vnější jaderné vrstvě (obr. 2A), což vede ke zhoršení optických vlastností sítnice. Nedávno demonstrovali Irina Solovei a spol.2) v elegantní studii, že u nočních savců je tato nevýhoda kompenzována unikátní přestavbou jader tyčinek.
Unikátní „invertovaná“ architektura jádra tyčinek myší sítnice
 Všechny myší buňky, včetně buněk sítnice, jakými jsou např. gangliové, bipolární buňky nebo čípky, mají chromatin v jádře uspořádán běžným způsobem: heterochromatin tvoří několik chromocenter, která jsou přimknutá k okraji jádra nebo k jadérku, a prstenec na periferii jádra; euchromatin je uvnitř jádra. Většina přepisovaných genů je tedy lokalizována uvnitř jádra, intenzivní transkripce však probíhá také na jeho periferii v blízkosti jaderných pórů.
Všechny myší buňky, včetně buněk sítnice, jakými jsou např. gangliové, bipolární buňky nebo čípky, mají chromatin v jádře uspořádán běžným způsobem: heterochromatin tvoří několik chromocenter, která jsou přimknutá k okraji jádra nebo k jadérku, a prstenec na periferii jádra; euchromatin je uvnitř jádra. Většina přepisovaných genů je tedy lokalizována uvnitř jádra, intenzivní transkripce však probíhá také na jeho periferii v blízkosti jaderných pórů.
V jádrech tyčinek je tomu přesně naopak: heterochromatin tvoří jedno velké, kompaktní chromocentrum ve středu jádra; euchromatin je na periferii (obr. 2). Kódující geny jsou tedy na periferii, nehledě na jejich polohu na chromozomu a transkripční status. Na periferii jsou lokalizovány jak geny aktivně přepisované (geny kódující provozní proteiny3) i proteiny specifické pro tyčinky), tak i geny, které v tyčinkách přepisovány nejsou. Také jadérko, transkripční faktory, vznikající primární transkripty a aparát nutný k sestřihu primárních transkriptů jsou na periferii jádra.
 Je zajímavé, že se tato specifická organizace jádra ustavuje až během konečné fáze diferenciace tyčinky (obr. 3). Jádra tyčinek novorozených myší mají normální strukturu. Během postnatálního vývoje se heterochromatin postupně přeskupuje do středu jádra, přičemž jednotlivá chromocentra splývají v jediné centrální. Euchromatin a jadérko se naopak přesouvají na periferii. Celé jádro se zmenšuje a zakulacuje. Celý proces „inverze“ jaderné organizace je završen přibližně ve věku jednoho měsíce. V tomto kontextu není bez zajímavosti poznamenat, že zrakové schopnosti myší se plně rozvíjejí až okolo 4 týdnů věku.
Je zajímavé, že se tato specifická organizace jádra ustavuje až během konečné fáze diferenciace tyčinky (obr. 3). Jádra tyčinek novorozených myší mají normální strukturu. Během postnatálního vývoje se heterochromatin postupně přeskupuje do středu jádra, přičemž jednotlivá chromocentra splývají v jediné centrální. Euchromatin a jadérko se naopak přesouvají na periferii. Celé jádro se zmenšuje a zakulacuje. Celý proces „inverze“ jaderné organizace je završen přibližně ve věku jednoho měsíce. V tomto kontextu není bez zajímavosti poznamenat, že zrakové schopnosti myší se plně rozvíjejí až okolo 4 týdnů věku.
Invertovaná architektura jádra je společná savcům s noční aktivitou
Proč mají právě tyčinky unikátní stavbu jádra? Odpověď na tuto otázku naznačila široká srovnávací analýza.
Autoři výše zmíněné studie2) vyšetřili okolo 40 druhů savců a nalezli jednoznačnou korelaci mezi stavbou tyčinek a způsobem života. Noční savci mají invertovanou architekturu jádra, zatímco savci s denní aktivitou mají jádro tyčinek organizováno běžným způsobem (obr. 4). Přechodný typ organizace jádra byl nalezen pouze u skotu. Ještě těsněji stavba jádra koreluje s ostatními adaptacemi sítnice na denní, respektive noční vidění, tedy s absolutní hustotou tyčinek a podílem čípků.
 Dosud ne zcela zodpovězenou otázkou zůstává, zda invertovaná architektura jádra vznikla v evoluci několikrát nezávisle u různých skupin savců s noční aktivitou, nebo vznikla jen jednou a byla několikrát ztracena u savců s aktivitou denní. Pravděpodobnější se zdá být druhá varianta. Savci byli zřejmě až do konce druhohor nočními tvory a teprve po vymření dinosaurů prošli adaptivní radiací spojenou s druhotným přechodem mnoha nepříbuzných skupin savců k denní aktivitě. Zásadní adaptace na noční vidění tedy pravděpodobně vznikly v rané evoluci savců (viz výše).
Dosud ne zcela zodpovězenou otázkou zůstává, zda invertovaná architektura jádra vznikla v evoluci několikrát nezávisle u různých skupin savců s noční aktivitou, nebo vznikla jen jednou a byla několikrát ztracena u savců s aktivitou denní. Pravděpodobnější se zdá být druhá varianta. Savci byli zřejmě až do konce druhohor nočními tvory a teprve po vymření dinosaurů prošli adaptivní radiací spojenou s druhotným přechodem mnoha nepříbuzných skupin savců k denní aktivitě. Zásadní adaptace na noční vidění tedy pravděpodobně vznikly v rané evoluci savců (viz výše).
Navíc se zdá, že organizace jádra je nesmírně konzervativním znakem, jehož modifikace je podstatně složitější než modifikace poměrného zastoupení jednotlivých fotoreceptorů. Například morče nebo myš nilská vykazují, na rozdíl od svých příbuzných, denní aktivitu. Zatímco mozaika jejich fotoreceptorů je již plně přizpůsobena dennímu vidění (nízká hustota tyčinek a vysoký podíl čípků), jádra jejich tyčinek si podržela invertovanou organizaci.
Ilustrativní je také příklad opačný. Vyšší primáti jsou denními tvory, jedinou výjimkou jsou mirikiny. Přestože jsou tyto jihoamerické opice silně přizpůsobeny k nočnímu životu, sdílejí s ostatními vyššími primáty konvenční stavbu jádra. Našly však alternativní způsob, jak vylepšit optické vlastnosti vnější jaderné vrstvy sítnice: uprostřed jádra každého fotoreceptoru je umístěno velké jadérko. Unikátní architektura jádra tyčinek tedy patrně vznikla v evoluci jen jednou, zatímco běžná stavba jádra byla několikrát znovu získána u různých fylogeneticky nepříbuzných skupin savců, kteří se plně adaptovali na denní aktivitu.
Invertované jádro funguje jako sběrná čočka soustřeďující světlo na vnitřní segmenty tyčinek
Jakým způsobem invertovaná architektura jádra přispívá k nočnímu vidění? Optické experimenty a počítačové simulace dokládají, že optimalizuje přenos světla přes vnější jadernou vrstvu sítnice (obr. 5).
 Malé jádro s centrálně lokalizovaným, vysoce kondenzovaným heterochromatinem má specifické optické vlastnosti. Index lomu centrálního heterochromatinu je vyšší (pravděpodobně díky vysoké koncentraci proteinů) než index lomu periferního euchromatinu. Celé jádro se pak chová jako konvexní čočka (spojka), která soustředí procházející světlo na vnitřní segmenty tyčinek. Naproti tomu jádro s konvenční architekturou chromatinu je opticky podstatně heterogennější. Vyšší index lomu je zde asociován s jednotlivými chromocentry a s periferním prstencem heterochromatinu. Při průchodu světla konvenčním jádrem proto dochází k vyššímu rozptylu světla.
Malé jádro s centrálně lokalizovaným, vysoce kondenzovaným heterochromatinem má specifické optické vlastnosti. Index lomu centrálního heterochromatinu je vyšší (pravděpodobně díky vysoké koncentraci proteinů) než index lomu periferního euchromatinu. Celé jádro se pak chová jako konvexní čočka (spojka), která soustředí procházející světlo na vnitřní segmenty tyčinek. Naproti tomu jádro s konvenční architekturou chromatinu je opticky podstatně heterogennější. Vyšší index lomu je zde asociován s jednotlivými chromocentry a s periferním prstencem heterochromatinu. Při průchodu světla konvenčním jádrem proto dochází k vyššímu rozptylu světla.
Vnější jaderná vrstva sítnice je tvořena několika vrstvami jader fotoreceptorů (obr. 1A). Díky větší hustotě tyčinek je těchto vrstev u nočních savců více (6–14) než u savců denních (≤7). Jednotlivá jádra jsou uspořádána nad sebou do sloupců orientovaných ve směru šíření světla. Při průchodu světla sloupcem jader s konvenční architekturou chromatinu dochází k významnému rozptylu a světlo se šíří i do sousedních sloupců. Naopak při průchodu světla sloupcem jader s invertovanou architekturou je rozptyl minimalizován a světlo se mezi sousedními sloupci nemísí. Unikátní struktura jádra tyčinek tedy zlepšuje optické vlastnosti sítnice.
Živé světlovody sítnice aneb Optika, o které se v učebnicích zatím nedozvíme
Oko obratlovců je na míle vzdáleno od „inženýrsky“ dokonalého řešení. Světlo musí projít celou sítnicí, aby dosáhlo vnějších, světločivých segmentů fotoreceptorů. Tento suboptimální design daný evoluční historií oka je kompenzován řadou důmyslných adaptací.
Efektivní průchod světla sítnicí savců se zdá být zajištěn hned několika strukturními elementy sítnice, které fungují jako „živé světlovody“ a minimalizují rozptyl světla. V brilantní studii demonstrovali Kristian Franze a kolektiv4), že jako optická vlákna fungují Müllerovy gliové buňky. Tyto gliové buňky začínají nálevkovitým rozšířením na vnitřní straně sítnice a radiálně (tj. ve směru šíření světla) ji celou prostupují. Index lomu Müllerových buněk je vyšší než index lomu okolní tkáně. Navíc se index lomu mění podél radiální osy gliové buňky: v místě „nálevkovité nožky“ je nejnižší a téměř shodný s indexem lomu sklivce – jak se buňka zužuje, její index lomu roste. Nálevkovité nožky pokrývají celý vnitřní povrch sítnice. Tyto vlastnosti činí z Müllerových buněk téměř dokonalé světlovody minimalizující nejen rozptyl, ale také lom světla na přechodu mezi sklivcem a sítnicí.
Nicméně ve vnější jaderné vrstvě sítnice se Müllerovy buňky rozvětvují okolo jader fotoreceptorů. Je tedy pravděpodobné, že efektivně vedou světlo od vnitřního okraje sítnice k vnější jaderné vrstvě, kde roli světlovodu přebírají „mikročočky“ tvořené sloupci jader tyčinek, které světlo přenášejí na vnitřní segmenty fotoreceptorů. Vnitřní segmenty fotoreceptorů jsou napěchovány mitochondriemi orientovanými podél jejich osy a samy tak fungují jako efektivní světlovody kanalizující průchozí světlo na vnější, světločivé segmenty fotoreceptorů.
Poznámky
1) Peichl L.: Diversity of mammalian photoreceptor properties: Adaptations to habitat and lifestyle? Anatomical Record A, 287A(1), 1001–1012, 2005.
2) Solovei I. et al.: Nuclear architecture of rod photoreceptor cells adapts to vision in mammalian evolution. Cell, 137(2), 356–368, 2009.
3) Provozní proteiny (housekeeping proteins) jsou přepisovány ve všech buňkách. Jde např. o enzymy základních metabolických drah, hlavní strukturní proteiny cytoskeletu a chromozomů, základní proteiny endoplazmatického retikula a Golgiho aparátu, ribozomální proteiny atd.
4) Kristian Franze et al.: Müller cells are living optical fibers in the vertebrate retina. PNAS, 104(20), 8287–8292, 2007.
Eukaryotní jádro
Jádro je informačním centrem eukaryotní buňky, ve formě DNA obsahuje valnou většinu její genetické výbavy. Jádro typické savčí buňky má průměr menší než 10 μm a obsahuje DNA o celkové délce přibližně 2 metry. DNA je pomocí histonů a dalších specializovaných proteinů sbalena takovým způsobem, který jí umožňuje interakci se všemi proteiny potřebnými pro její transkripci, replikaci a reparaci. Komplex DNA, RNA a proteinů vytvářející chromozomy nazýváme chromatin. V době buněčného dělení jsou chromozomy v maximálně kondenzovaném stavu, v interfázi mají mnohem rozvolněnější strukturu. Obecně řečeno, oblasti chromozomu, které jsou transkribovány, mají rozvolněnější strukturu, nepřepisované oblasti jsou kompaktnější, více spiralizované. V chromozomech se vyskytují dva základní typy chromatinu. Kompaktnější formou chromatinu je heterochromatin, který obvykle tvoří více než 10 % interfázního chromatinu a je transkripčně neaktivní. Zbytek chromatinu, euchromatin, je spiralizován méně a nachází se v něm většina genů. U typické diferencované buňky je aktivně transkribováno přibližně 10 % chromatinu. Heterochromatin a euchromatin jsou prostorově odděleny nejen na chromozomu, ale i v jádře. Heterochromatin se typicky koncentruje na periferii jádra a poblíž jadérka, kde vytváří shluky (chromocentra), euchromatin je lokalizován spíše ve středu jádra. Takovéto uspořádání je téměř univerzální, výjimku tvoří jen některé jednobuněčné organismy (např. obrněnky, některé kvasinky, makronukleus nálevníků). Obecně se soudí, že je tato organizace jádra optimální pro prostorovou separaci transkripčně aktivního a neaktivního chromatinu a pro prostorové interakce mezi geny; obé pravděpodobně hraje důležitou roli v regulaci transkripce. Vzhledem k funkčnímu významu a evoluční stabilitě této jaderné organizace bylo velkým překvapením, že se prostorové uspořádání chromatinu v jádrech tyčinek myší sítnice zásadně odlišuje od výše uvedeného vzoru.
Ke stažení
 článek ve formátu pdf [735,73 kB]
článek ve formátu pdf [735,73 kB]
O autorovi
Pavel Němec
Doporučujeme

Rostliny vyprávějí o lidech

O prázdnech v nás