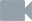Ploutvenky
 Do kmene ploutvenek (Chaetognatha) řadíme asi stovku druhů drobných mořských živočichů. Vyznačují se velmi neobvyklou stavbou těla, která jako by spojovala znaky různých evolučních větví živočišné říše. Problémem příbuzenských vztahů této skupiny se proto zabývala řada významných biologů, vzpomenout můžeme např. Ch. Darwina, A. Huxleyho, F. Leydiga, I. I. Mečnikova či A. O. Kowalewského. Ploutvenky byly postupně kladeny do okolí nejrůznějších živočišných skupin, například hlístic, vrtejšů, měkkýšů, korýšů, či dokonce strunatců. Přestože v posledních dvou desetiletích bylo o biologii ploutvenek získáno velké množství poznatků, nevíme stále nic bližšího o jejich skutečném postavení v systému živočichů ani o evoluci celé skupiny. Řešení problému je o to obtížnější, že embryonální vývoj je velmi neobvyklý a nebyl dosud studován novějšími technikami vývojové biologie. Chybí rovněž pravé larvální stadium, které by mohlo poskytnout jisté vodítko. Určitý pokrok nabízejí techniky molekulární biologie a studium fosilií. Díky zvláštní stavbě těla a neobvyklým smyslovým orgánům se ploutvenky dostávají rovněž do zájmové sféry fyziologů a buněčných biologů. Některé orgány se vyznačují značnou dokonalostí (nervová soustava), jiné mají naopak velmi jednoduchou stavbu (trávicí soustava) nebo vůbec chybějí (oběhová a vylučovací soustava).
Do kmene ploutvenek (Chaetognatha) řadíme asi stovku druhů drobných mořských živočichů. Vyznačují se velmi neobvyklou stavbou těla, která jako by spojovala znaky různých evolučních větví živočišné říše. Problémem příbuzenských vztahů této skupiny se proto zabývala řada významných biologů, vzpomenout můžeme např. Ch. Darwina, A. Huxleyho, F. Leydiga, I. I. Mečnikova či A. O. Kowalewského. Ploutvenky byly postupně kladeny do okolí nejrůznějších živočišných skupin, například hlístic, vrtejšů, měkkýšů, korýšů, či dokonce strunatců. Přestože v posledních dvou desetiletích bylo o biologii ploutvenek získáno velké množství poznatků, nevíme stále nic bližšího o jejich skutečném postavení v systému živočichů ani o evoluci celé skupiny. Řešení problému je o to obtížnější, že embryonální vývoj je velmi neobvyklý a nebyl dosud studován novějšími technikami vývojové biologie. Chybí rovněž pravé larvální stadium, které by mohlo poskytnout jisté vodítko. Určitý pokrok nabízejí techniky molekulární biologie a studium fosilií. Díky zvláštní stavbě těla a neobvyklým smyslovým orgánům se ploutvenky dostávají rovněž do zájmové sféry fyziologů a buněčných biologů. Některé orgány se vyznačují značnou dokonalostí (nervová soustava), jiné mají naopak velmi jednoduchou stavbu (trávicí soustava) nebo vůbec chybějí (oběhová a vylučovací soustava).
Základní schéma stavby ploutvenek
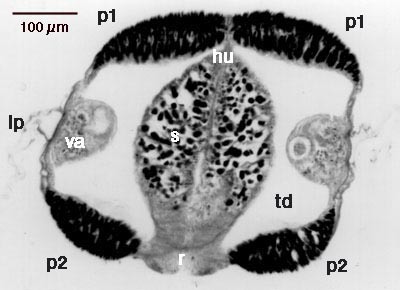
Průsvitné šípovité tělo ploutvenky je po stranách opatřeno jedním nebo dvěma páry vodorovných ploutviček, vyztužených chitinózními paprsky. Na zádi vyniká nepárová ocasní ploutev. Na těle můžeme rozeznat tři části (hlavu, trup a ocasní oddíl), které jsou anatomicky odděleny přepážkami. Ocasní oddíl je navíc rozdělen další přepážkou podélně (obrázek 1). Délka ploutvenek se pohybuje v rozmezí 0,2 – 12 cm.
Na těle můžeme rozeznat tři části (hlavu, trup a ocasní oddíl), které jsou anatomicky odděleny přepážkami. Ocasní oddíl je navíc rozdělen další přepážkou podélně (obrázek 1). Délka ploutvenek se pohybuje v rozmezí 0,2 – 12 cm.
Zploštělá hlava s párem očí nese několik párů tuhých chitinózních útvarů, které slouží k příjmu potravy. Jsou to mohutné svěrací ostny a mnohem menší „zoubky“ (obrázek 2, obrázek 13 a obrázek 22). Ze zadní části hlavy vyrůstá unikátní elastický kožní záhyb, který může podle potřeby překrýt celou hlavu jako kápě (obrázek 2, obrázek 13). Pohyb záhybu je usnadněn výměšky žláznaté tkáně, která tvoří jeho vnitřní povrch. Přehrnutá „kápě“ chrání před poškozením celý aparát sloužící k lovu kořisti. Zároveň jej tiskne k povrchu hlavy tak, aby nebrzdil pohyb těla.
Ploutvenky jsou rychlí a vytrvalí plavci
Značná pohyblivost podpořila vývoj specializované oporné soustavy a svalstva, které ve své stavbě spojuje znaky tak vzdálených skupin, jakými jsou hlístice, členovci a obratlovci.Největší část tělní stěny je tvořena tzv. primárním pohybovým svalstvem (obrázek 20), které se vyznačuje velmi komplikovanou stavbou. V těchto podélných, příčně pruhovaných svalech se pravidelně střídají dva typy svalových vláken, označované A a B (obrázek 17, obrázek 19). Pravidelné uspořádání vláken odráží způsob jejich vzniku. Protože ploutvenky rostou během celého života, vytvářejí se neustále na okrajích primárních svalů dva typy buněk, myoblasty A a B (obrázek 19, označené z). Oba typy myoblastů se dělí na konstantní počet buněk, které se mění na svalová vlákna příslušného typu. Ve svalu se pak pravidelně střídají skupiny vláken A a B. Výjimkou jsou primární svaly čeledi Spadellidae, složené jen z vláken typu A.
 Oba typy vláken se liší jak ultrastrukturou, tak biochemicky. U obou typů vytváří cytoplazmatická membrána hluboké vchlípeniny, které jsou u vláken B v těsném kontaktu se systémem sarkoplazmatického retikula (obrázek 3, obrázek 17, obrázek 18). Ultrastruktura vláken B se tak až zarážejícím způsobem blíží poměrům, jaké nalézáme ve svalových vláknech kosterního svalstva obratlovců. U těch se systém vchlípenin (tzv. tubulů T) a sarkoplazmatického retikula podílí na zdokonalení vztahu mezi podrážděním a stahem vlákna. Přes tuto podobnost má primární svalstvo ploutvenek mnohé jedinečné rysy, které se u obratlovců nevyskytují. 1) Stavbou kontraktilního aparátu svalových vláken a mechanickými vlastnostmi se primární svalovina blíží spíše nepřímým křídelním svalům hmyzu.
Oba typy vláken se liší jak ultrastrukturou, tak biochemicky. U obou typů vytváří cytoplazmatická membrána hluboké vchlípeniny, které jsou u vláken B v těsném kontaktu se systémem sarkoplazmatického retikula (obrázek 3, obrázek 17, obrázek 18). Ultrastruktura vláken B se tak až zarážejícím způsobem blíží poměrům, jaké nalézáme ve svalových vláknech kosterního svalstva obratlovců. U těch se systém vchlípenin (tzv. tubulů T) a sarkoplazmatického retikula podílí na zdokonalení vztahu mezi podrážděním a stahem vlákna. Přes tuto podobnost má primární svalstvo ploutvenek mnohé jedinečné rysy, které se u obratlovců nevyskytují. 1) Stavbou kontraktilního aparátu svalových vláken a mechanickými vlastnostmi se primární svalovina blíží spíše nepřímým křídelním svalům hmyzu.
Epitel kryjící povrch těla je u ploutvenek na rozdíl od většiny bezobratlých (a podobně jako u obratlovců) složen z mnoha vrstev. Novější studie potvrdily, že se mnohovrstevný epitel nevyskytuje na celém povrchu těla. Omezuje se na oblasti, které jsou podloženy svalstvem zabezpečujícím pohyb. To naznačuje, že se vícevrstevnatost krycího epitelu vyvinula teprve druhotně, v souvislosti se začleněním do oporného systému. Mnohovrstevný epitel by tedy měl fungovat spolu s bazální blánou, která jej podkládá, jako vnější oporný systém – exoskelet. Pro tyto závěry svědčí i výsledky studia elektronovým mikroskopem. Spodní vrstvy epitelu jsou totiž složeny z buněk, které do sebe zapadají četnými výběžky, což velmi zvyšuje soudržnost tkáně. Opornou funkci epitelu umocňuje tlak zvláštní tekutiny (hydroskeletu), která vyplňuje rozsáhlé dutiny uvnitř těla (obrázek 20, označeno td). Tlak hydroskeletu na stěny je regulován objemovými změnami dutin, které řídí výstelka specializovaných svalových buněk. Stabilní složení hydroskeletu zabezpečují specializované, gigantické střevní buňky, jež vyčnívají ze střeva do tělních dutin a podle potřeby hromadí odpadní látky.
V evoluci ploutvenek vznikly rozmanité mechanizmy, které zmenšují hustotu těla tak, aby bylo umožněno dlouhodobé splývání. U některých druhů je splývání podporováno již zmíněnými obrovskými střevními buňkami, které díky své nízké hustotě nadlehčují tělo. Podobná úloha je přisuzována zvláštní pokožce z velkých buněk, která často pokrývá část těla (obrázek 22). V některých případech je dokonce redukována i svalová hmota. Přesto má tělo většiny ploutvenek poněkud větší hustotu než mořská voda, a proto ve vodním sloupci pomalu klesá. Tato tendence je kompenzována periodickými pohyby, které zabezpečuje primární pohybové svalstvo. Střídavá aktivita dorzálních a ventrálních primárních pohybových svalů vede k sérii bleskurychlých záškubů, jimiž je tělo doslova „vystřeleno“ nahoru. Celý pohyb je pasivně usměrňován ploutvičkami. Podobné chování se uplatňuje také při lovu potravy.
Potrava ploutvenek
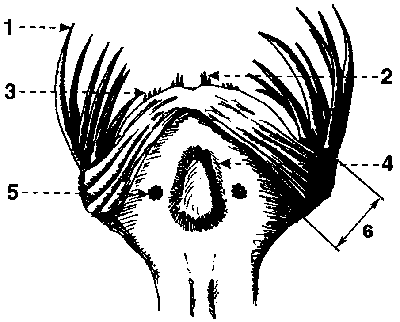
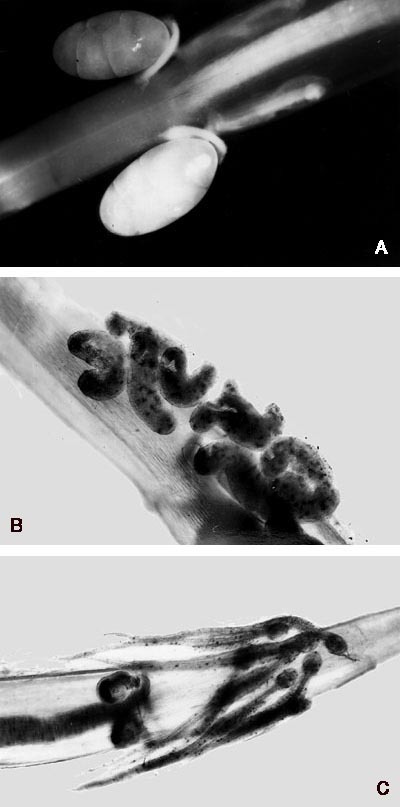
Ploutvenky se živí zejména drobnými korýši, prvoky a jinými ploutvenkami, v menší míře také vajíčky a larvami ryb. Kořist zachycují svěracími ostny, jimiž je vyzbrojena hlava ploutvenky. Ostny jsou velmi pevné, a přitom i pružné útvary: jsou tvořeny polysacharidem chitinem s příměsí zinku, špičky jsou z oxidu křemičitého.Chování polapené kořisti vedlo již dříve mnohé autory k domněnce, že ploutvenky používají při lovu nějaký jed. Teprve r. 1988 prokázal americký badatel E. V. Thuesen, že jde o tetrodotoxin, velmi účinnou látku, která je neurotoxinem blokujícím sodné kanálky na povrchu nervových buněk (vzorec této látky je na obrázku 4). Pro člověka je smrtelný již v dávce 10 mg/kg tělesné hmotnosti. Čistý tetrodotoxin je velmi stabilní, ve vodě nerozpustná sloučenina, která se rozkládá teprve působením silných kyselin a zásad. Je zajímavé, že tetrodotoxin je znám také u mnoha jiných, většinou mořských živočichů. Nejprve byl popsán u čtverzubcovitých ryb, podle jejichž latinského označení (Tetraodontidae) byl pojmenován. Později byl nalezen i u dalších skupin: plžů, hlavonožců, korýšů, ostnokožců a obojživelníků. Podrobný výzkum vedl k překvapivému zjištění: jed neprodukují živočichové samotní, nýbrž bakterie rodu Vibrio, zejména druh V. alginolyticus. Znamená to, že živočichové, u kterých byl tetrodotoxin nalezen, jsou jedovatí druhotně. Nejčastěji se buď oni sami, nebo jejich kořist živí bakteriemi, které produkují toxin. V důsledku své mimořádné stability se tetrodotoxin nakonec hromadí v jejich tělech. Ploutvenky a chobotnice Hapalochlaena maculosa si bakterie „chovají“ a využívají tetrodotoxin při lovu kořisti. Symbionti ploutvenek žijí pravděpodobně v hlavové části hostitele. Dosud přesně nevíme, které orgány slouží k přechovávání symbiontů ani kde ploutvenky shromažďují jed a jak jej nakonec vpraví do kořisti (podrobněji obrázek 13). Protože je jed produkován v nepatrném množství, mohl být identifikován teprve až velmi citlivými chromatografickými technikami. Ploutvenky tak sice nejsou nebezpečné lidem, mohou však roznášet jedovaté bakterie v mořských ekosystémech, neboť se hojně vyskytují v povrchových vrstvách všech oceánů.
Při orientaci v prostoru využívají ploutvenky řadu smyslových orgánů
Některé smyslové orgány ploutvenek mají velmi neobvyklou strukturu a funkci. Tomu odpovídá pokročilá stavba nervové soustavy. Její hmota je soustředěna do několika uzlin (ganglií), které jsou převážně v hlavové části. Nejvíce poznatků máme o největší, hlavní uzlině, uložené v přední části hlavy. Její povrch je na rozdíl od ostatních ganglií kryt dvouvrstevným obalem, který se svou stavbou velmi podobá myelinové pochvě nervových buněk obratlovců. Do svrchní části hlavní uzliny se zanořuje retrocerebrální orgán, tvořený dvěma bohatě rozvětvenými chodbami, které ústí společným vývodem přímo na povrch těla. V uzlině pronikají do těchto chodeb zvláštní nervové buňky. Funkce orgánu je nejasná, předpokládá se však, že je v něm provázána endokrinní činnost s určitými typy smyslového vnímání. Podobné struktury se vyskytují také u dalších skupin bezobratlých (např. pásnic a vířníků), ale ani zde není jejich význam příliš jasný. Hlavní uzlina vysílá nervy také k páru očí, které leží uprostřed svrchní strany hlavy. Oči ploutvenek se vyznačují rozmanitou stavbou. Příkladem nám bude nejprve druh Flaccisagitta hexaptera, který se vyskytuje v povrchové vrstvě oceánu. Jeho oko obsahuje více než 400 senzorických buněk, jež jsou rozděleny do tří skupin velkou pigmentovou buňkou ve tvaru T (obrázek 9). Světločivé části zrakových buněk jsou odvráceny od dopadajícího světla (obrázek 12). To znamená, že světlo musí, dříve než je zachyceno, projít tělem zrakové buňky. Výsledky prvních studií vedly k názoru, že světločivé ultrastruktury těchto buněk (rhabdom) jsou tvořeny svazkem tyčinkovitých výběžků, které obsahují pigment rodopsin. Takové uspořádání by vcelku odpovídalo situaci, jakou nacházíme u řady jiných skupin živočichů.
Hlavní uzlina vysílá nervy také k páru očí, které leží uprostřed svrchní strany hlavy. Oči ploutvenek se vyznačují rozmanitou stavbou. Příkladem nám bude nejprve druh Flaccisagitta hexaptera, který se vyskytuje v povrchové vrstvě oceánu. Jeho oko obsahuje více než 400 senzorických buněk, jež jsou rozděleny do tří skupin velkou pigmentovou buňkou ve tvaru T (obrázek 9). Světločivé části zrakových buněk jsou odvráceny od dopadajícího světla (obrázek 12). To znamená, že světlo musí, dříve než je zachyceno, projít tělem zrakové buňky. Výsledky prvních studií vedly k názoru, že světločivé ultrastruktury těchto buněk (rhabdom) jsou tvořeny svazkem tyčinkovitých výběžků, které obsahují pigment rodopsin. Takové uspořádání by vcelku odpovídalo situaci, jakou nacházíme u řady jiných skupin živočichů.
 Revizní studie na konci osmdesátých let však původní názory nepotvrdila. Odhalila bizarní, složitou lamelární strukturu, proděravělou navíc četnými póry (viz obrázek 12). Japonský badatel T. Goto se později v rozsáhlé práci zabýval stavbou očí u několika dalších zástupců čeledi Sagittidae. Zjistil, že stavba očí se u jednotlivých druhů liší podle hloubky, v níž určitý druh žije. Se vzrůstající hloubkou se centrální pigmentová buňka zmenšuje (a u hlubinných druhů i mizí), kdežto plocha obsazená zrakovými buňkami se zvětšuje. V očích může vzniknout více oddělených skupin zrakových buněk. Má se za to, že takové oči by mohly pracovat podobně jako složené oči členovců (například korýšů).
Revizní studie na konci osmdesátých let však původní názory nepotvrdila. Odhalila bizarní, složitou lamelární strukturu, proděravělou navíc četnými póry (viz obrázek 12). Japonský badatel T. Goto se později v rozsáhlé práci zabýval stavbou očí u několika dalších zástupců čeledi Sagittidae. Zjistil, že stavba očí se u jednotlivých druhů liší podle hloubky, v níž určitý druh žije. Se vzrůstající hloubkou se centrální pigmentová buňka zmenšuje (a u hlubinných druhů i mizí), kdežto plocha obsazená zrakovými buňkami se zvětšuje. V očích může vzniknout více oddělených skupin zrakových buněk. Má se za to, že takové oči by mohly pracovat podobně jako složené oči členovců (například korýšů).
Zajímavá stavba očí u čeledi Sagittidae dala podnět k podobně zaměřenému výzkumu u dalších ploutvenek. Bylo zjištěno, že vypouklé oči rodu Eukrohnia (Eukrohniidae) se svou stavbou dosti podobají složeným očím členovců. Jsou tvořeny až stovkou drobných oček, přičemž světločivé struktury zrakových buněk jsou přivráceny ke zdroji světla (viz obrázek 10 a obrázek 11). Čočky jednotlivých oček se nacházejí těsně pod povrchem těla, kryty jen nepatrnou blankou. Světločivé struktury těchto ploutvenek leží, na rozdíl od obdobných struktur členovců, velmi blízko povrchu těla – lemují čočky po stranách (viz obrázek 11). Experimenty naznačují, že i přes rozdíly ve stavbě fungují oči rodu Eukrohnia stejně jako složené oči členovců.
Původní typ stavby očí u ploutvenek reprezentují podle Francouze F. Ducreta oči druhu Flaccisagitta hexaptera. Je zajímavé, že velmi podobná stavba byla nedávno popsána také u bentických (u dna žijících) ploutvenek čeledi Spadellidae, které jsou i na základě dalších indicií (nemají např. svalová vlákna typu B) pokládány za nejpůvodnější žijící formu kmene. Teprve mnohem později se vyvinuly „složené“ oči, jak je známe u rodu Eukrohnia.
Soudí se, že při orientaci v prostoru hrají oči ploutvenek jen podřadnou úlohu. Větší význam je v tomto směru přisuzován rozmanitým obrveným receptorům na povrchu těla, zejména tzv. obrveným políčkům. Jsou to skupiny brv dlouhých až několik stovek mikrometrů, pravidelně rozmístěné po celém povrchu těla včetně ploutviček. Na trupu a ocasní části lze odlišit řady větších políček, které probíhají paralelně s podélnou osou těla, a na ně kolmé řady menších políček. U druhu Flaccisagitta enflata bylo na těle dospělce dlouhého 18 mm napočítáno více než 600 těchto struktur. Experimentálně bylo zjištěno, že jde o jistou obdobu proudového orgánu u ryb: políčka zachycují širokou škálu vibrací, jejichž zdroj se nachází poblíž těla. Studiem útoků ploutvenek na mechanické vibrátorky bylo také potvrzeno, že zmíněná políčka jsou nejcitlivější na kmitočty vyvolané klanonožci (Copepoda). Tato skupina drobných planktonických korýšů tvoří hlavní součást potravy ploutvenek.
 Význam dalších typů obrvených receptorů je dosud nejasný. Do souvislosti s rozmnožováním bývá dávána tzv. corona ciliata, nápadný útvar na svrchní straně těla – na rozhraní hlavy a trupu. Je tvořen oválným žláznatým políčkem, lemovaným na obvodu zvláštním typem brv (viz obrázek 1 a obrázek 2). U rodu Spadella se zjistilo, že žláznaté buňky políčka produkují v době rozmnožování zvláštní sekret, který stéká k vyústění samičí pohlavní soustavy. Po této stopě se pak při oplození pohybují spermie.
Význam dalších typů obrvených receptorů je dosud nejasný. Do souvislosti s rozmnožováním bývá dávána tzv. corona ciliata, nápadný útvar na svrchní straně těla – na rozhraní hlavy a trupu. Je tvořen oválným žláznatým políčkem, lemovaným na obvodu zvláštním typem brv (viz obrázek 1 a obrázek 2). U rodu Spadella se zjistilo, že žláznaté buňky políčka produkují v době rozmnožování zvláštní sekret, který stéká k vyústění samičí pohlavní soustavy. Po této stopě se pak při oplození pohybují spermie.
Ploutvenky jsou obojetníci
U jednoho individua najdeme jak samčí, tak samičí pohlavní žlázy. Přesto se samooplození vyskytuje jen výjimečně, protože spermie dozrávají dříve. Oba typy pohlavních žláz se vyznačují ojedinělou stavbou. Párové vaječníky jsou uloženy v zadní části trupu. Do jejich dutiny vyčnívají vajíčka, nesená zvláštní dvoubuněčnou stopkou (viz obrázek 5). Je zajímavé, že vaječník je zcela uzavřený a nemá spojení se slepou chodbou (ovispermaduktem), která jej po celé délce lemuje a slouží jednak k přístupu spermií, jednak k uvolňování oplozených vajíček. Při vlastním oplození musí spermie patrně proniknout stěnou vaječníku a buňkami, které tvoří stopku vajíčka. Po oplození stopka degeneruje a vajíčka se protržením stěny vaječníku dostávají do vývodné chodby, a odtud do vody. Některé hlubinné druhy „vychovávají“ potomstvo ve dvou zvláštních váčcích uložených po stranách těla, v místech vyústění samičí pohlavní soustavy. Tyto pozoruhodné útvary popsal jako první japonský badatel M. Terazaki (viz obrázek 14, obrázek 15 a obrázek16).
Párové vaječníky jsou uloženy v zadní části trupu. Do jejich dutiny vyčnívají vajíčka, nesená zvláštní dvoubuněčnou stopkou (viz obrázek 5). Je zajímavé, že vaječník je zcela uzavřený a nemá spojení se slepou chodbou (ovispermaduktem), která jej po celé délce lemuje a slouží jednak k přístupu spermií, jednak k uvolňování oplozených vajíček. Při vlastním oplození musí spermie patrně proniknout stěnou vaječníku a buňkami, které tvoří stopku vajíčka. Po oplození stopka degeneruje a vajíčka se protržením stěny vaječníku dostávají do vývodné chodby, a odtud do vody. Některé hlubinné druhy „vychovávají“ potomstvo ve dvou zvláštních váčcích uložených po stranách těla, v místech vyústění samičí pohlavní soustavy. Tyto pozoruhodné útvary popsal jako první japonský badatel M. Terazaki (viz obrázek 14, obrázek 15 a obrázek16).
Párová varlata jsou umístěna v ocasním oddílu. V období pohlavního zrání stěna varlat praská a do tekutiny vyplňující ocasní oddíl se uvolňují tzv. spermatogonie, což jsou buňky, které dalším dělením dávají vzniknout pohlavním buňkám. Víření obrveného epitelu na stěnách ocasní dutiny vyvolává neustálý pohyb zrajících pohlavních buněk v tekutině. Do vývodných cest se dostávají teprve zralé spermie. Vývodné chodby jsou slepé, končí ve zvláštních zásobních pouzdrech, tzv. vesiculae seminales (viz obrázek 1). Teprve po prasknutí pouzder se uvolňují do vody shluky spermií slepené želatinózní hmotou.
Ve vláknitých spermiích ploutvenek nalezneme stejné ultrastruktury jako u jiných živočichů, jsou však neobvykle uspořádány. U klasického typu vláknité, resp. bičíkaté spermie obsahuje hlavička jádro, za nímž leží v bázi bičíku jedna nebo dvě centrioly. Pohyb bičíku je vyvolán tzv. osním vláknem bičíku (axonematem), které navazuje na centrioly. Ve spermiích ploutvenek najdeme centriolu již v přední části hlavičky, navazující axonema probíhá celou spermií až do bičíku. Jádro leží teprve v zadní části spermie. Vzniká tak dojem, že celá spermie je tvořena pouze „bičíkem“ (viz obrázek 6 vlevo). Zvláštní stavba spermií patrně souvisí s neobvyklým průběhem oplození popsaným výše.
Atypická stavba pohlavních orgánů vede autory článku k názoru, že u předků ploutvenek měly pohlavní orgány odlišné vývodné cesty, než nacházíme u současných forem. Tyto primární vývody byly patrně ztraceny v průběhu evoluce při drastické změně celkové stavby těla. Dnešní vývody jsou pravděpodobně jen druhotnými strukturami.
Z vajíček se líhne larvální stadium, které nepřijímá potravu a postrádá některé znaky dospělců. Chybí například aparát k lovu kořisti, oči, postranní ploutvičky a řitní otvor (viz obrázek 6 vpravo). Mladé ploutvenky se velmi rychle vyvíjejí, takže se již během několika dnů téměř neliší od dospělců. Nejpozději se tvoří pohlavní vývody a přepážka, která odděluje samčí a samičí pohlavní žlázy a zároveň i ocasní oddíl od trupu. Je zajímavé, že u některých druhů (Adhesisagitta hispida) je larvální stadium značně zkráceno a vylíhnuté ploutvenky jsou velmi podobné dospělcům.
Většina ploutvenek se vyznačuje ohromnými areály rozšíření
Ploutvenky, podobně jako mnoho dalších mořských organizmů žijících v planktonu, obývají rozsáhlé oblasti. Areály některých druhů mohou zahrnovat i několik oceánů. Schopnost šíření je podmíněna zejména tolerancí ke změnám faktorů mořského prostředí a produkcí velkého množství vajíček, která jsou roznášena mořskými proudy. Největší druhové bohatství nalezneme v tropech a subtropech. Většina zástupců se vyskytuje v povrchových vrstvách otevřeného moře (takzvaném epipelagiálu), některé druhy jsou však vázány na oblast šelfu (neritikum). Pouze zástupci čeleďi Spadellidae se vyskytují u dna. Adaptací k takovému způsobu života jsou zvláštní příchytné orgány na ploutvičkách, které jsou nejlépe vyvinuty u rodu Paraspadella. U druhu P. gotoi jsou vybaveny vlastním svalstvem, podobají se končetinám a mohou udržovat tělo i ve vertikální poloze.Pelagické druhy vykonávají během dne – jako mnoho jiných planktonních skupin – vertikální migraci, a to v rozsahu několika stovek metrů. Příčiny tohoto jevu nebyly dosud uspokojivě vysvětleny. Současné hypotézy zdůrazňují hojnost potravy v menších hloubkách a zpomalení metabolizmu při potopení do větších hloubek. Během migrace se ploutvenky orientují převážně zrakem – jsou to totiž živočichové reagující pozitivně na světlo.
 Vzhledem k hojnému výskytu v planktonních společenstvech jsou ploutvenky důležitou součástí mořských potravních řetězců. Ze současných objevů však vyplývá, že v řadě dalších ekosystémů jsou ploutvenky dosud přehlíženy. Specializované druhy byly nedávno popsány z podmořských jeskyní či z nepatrných mezer, které se vytvářejí mezi zrnky v písčitých usazeninách. Druh Adhesisagitta hispida tráví zase většinu svého života na dně, přichycený výměšky zvláštních žláz.
Vzhledem k hojnému výskytu v planktonních společenstvech jsou ploutvenky důležitou součástí mořských potravních řetězců. Ze současných objevů však vyplývá, že v řadě dalších ekosystémů jsou ploutvenky dosud přehlíženy. Specializované druhy byly nedávno popsány z podmořských jeskyní či z nepatrných mezer, které se vytvářejí mezi zrnky v písčitých usazeninách. Druh Adhesisagitta hispida tráví zase většinu svého života na dně, přichycený výměšky zvláštních žláz.
Obrázek 7 ukazuje na několika vybraných zástupcích poměrně velkou tvarovou rozmanitost ploutvenek. Přesto je systematika této skupiny stále dosti problematická, a to zejména na nižších úrovních. Příčinou je především proměnlivost řady druhů. Je pozoruhodné, že různé formy jednoho druhu jsou morfologicky zřetelně odlišeny. Tyto druhové varianty můžeme nalézt nejen v geograficky vzdálených částech areálu, ale také na stejném místě v různých hloubkách, či dokonce ročních obdobích. Takové formy byly dříve běžně popisovány jako nové druhy. Teprve v poslední době dochází k žádoucí revizi těchto „druhů“, a to na základě analýz izoenzymů a sekvencí vybraných genů. První výsledky ukazují, že v některých případech jde o klony, rozmnožující se partenogenezí.
Problém původu ploutvenek je dosud zcela otevřený
Pokročilá stavba některých orgánů, rysy společné se strunatci a konečně i průběh embryonálního vývoje vedly k tomu, že byly ploutvenky v poslední době víceméně stabilně pojímány jako izolovaná evoluční větev tzv. druhoústých živočichů. Do této velké skupiny jsou kromě ploutvenek řazeny ještě další kmeny: ostnokožci, polostrunatci a strunatci. Některé práce z nedávné doby takové zařazení velmi zpochybnily a vyvolaly opět rozsáhlé diskuse o systematickém postavení ploutvenek. Studium ultrastruktur útrobního svalstva a přilehlých tkání ukázalo, že vnitřní dutina pravděpodobně neodpovídá tzv. druhotné tělní dutině (coelomu). Znamenalo by to, že ploutvenky nepatří dokonce ani do skupiny živočichů s coelomem, což je velký konglomerát kmenů, který zahrnuje kromě druhoústých živočichů ještě další skupiny. Srovnání sekvence genu pro 18S ribozomovou RNA s touto sekvencí u jiných živočichů vedlo již k úplnému oživení některých starších hypotéz (např. Schneider 1886), které zdůrazňovaly jistou příbuznost ploutvenek s hlísticemi či jinými skupinami s nepravou tělní dutinou (pseudocoelomem). Naše představy o příbuzenských vztazích ploutvenek však zůstávají i nadále velmi nepřesné. S napětím se proto očekává studium sekvence genu pro 18S rRNA u širšího spektra živočichů s pseudocoelomem a prostudování embryonálního vývoje ploutvenek moderními metodami.
Kmen Chaetognatha v geologické historii
K objasnění evoluce ploutvenek by jistě přispělo studium jejich fosilií – z geologické historie máme však jen málo dokladů. Je to způsobeno zejména tím, že jde o organizmy bez pevné kostry nebo schránky, jejichž fosilizace je výjimečnou záležitostí. V obsáhlé paleontologické literatuře jsou popsány pouhé tři druhy živočichů s tělem bez pevné kostry, které byly nebo dosud jsou považovány za fosilní ploutvenky. Jde vždy o zcela zploštělé jedince, kteří se zachovali jen jako tenký organický film na vrstevních plochách v jílovitých břidlicích nebo prachovcích.Stratigraficky i historicky nejstarší fosilní nálezy pocházejí ze středního kambria Kanady, z tzv. burgesských břidlic. V šedočerných jílovitých břidlicích se zachovaly v neobyčejném množství a diverzitě různé organizmy, často s otisky měkkých částí (končetin apod.). Tyto výjimečně zachovalé fosilie poprvé zkoumal a popsal známý paleontolog C. D. Walcott. Ten také roku 1911 uveřejnil popis druhu Amiskwia sagittiformis, který přiřadil k ploutvenkám. Tento taxon byl však později řazen i do jiných skupin a dnes panuje názor, že o ploutvenku nejde. Amiskwia se považuje za pásnici. K tomu, že byla přiřazena k uvedenému kmeni, přispělo zachování některých vnitřních orgánů, jejichž umístění a morfologie je bližší právě pásnicím. Některé pásnice se navíc vyznačují tvarovou konvergencí s ploutvenkami.
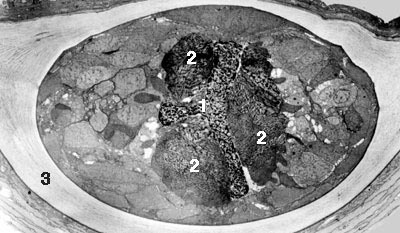
Titerina rokycanensis je druh, který byl popsán na základě tří exemplářů z českého spodního ordoviku. Byl nalezen v jílovitých břidlicích klabavského souvrství na severozápadním okraji Rokycan. Jediný kompletní jedinec je 4,9 mm dlouhý (viz obrázek 8A). I na úlomcích se však zachovaly zbytky orgánů, jejichž charakter a pozice je podobná jako u recentních ploutvenek (obrázek 8B). Celkově druh připomíná zástupce žijícího rodu Spadella.
Stratigraficky nejmladší, nicméně velmi problematické nálezy pocházejí ze severoamerického svrchního karbonu (pennsylvanianu), z oblasti Mazon Creek ve státě Illinois (USA). Našlo se zde několik druhů organizmů s červovitým tvarem těla, řazených mezi hlavatce (Priapulida), hlísty (Nematoda), pásnice (Nemertini) a ploutvenky. Nejistoty, které vznikají kolem taxonomického zařazení těchto fosilií, vycházejí z celkově nepříznivého zachování v konkrecích (nerostných útvarech jiného složení než okolní hornina), relativně hrubozrnných vzhledem k charakteru fosilních objektů. F. R. Schram odtud v roce 1973 popsal jako ploutvenky dva exempláře stínově zachovaných jedinců pod názvem Paucijaculum samamithion, které celkovým tvarem ploutvenky připomínají.
Pevné části těla ploutvenek, tj. zoubky a svěrací ostny, se mohou zachovat snáze, ale jsou zřejmě obecně přehlíženy. Dosud tak existuje jediný popis nálezů izolovaných svěracích ostnů. H. Szaniawski podrobil detailní analýze druh Prooneotodus? tenuis 2) ze svrchního kambria až spodního ordoviku, řazený do té doby mezi tzv. protokonodonty (organofosfatické mikrofosilie nejisté systematické příslušnosti, které mají tvar jednoduchých kuželovitých „zoubků“). V roce 1982 uveřejnil práci, v níž prokázal nápadnou morfologickou i mikrostrukturní podobnost tohoto druhu se svěracími ostny ploutvenek.
Žadný výše uvedený fosilní druh tedy není hodnocen jako průkazný zástupce ploutvenek. Nejistoty taxonomického zařazení je nutné přičíst především ztrátě některých důležitých znaků při fosilizaci. Velmi zavádějící však mohou být také snahy o nalezení vnitřních struktur dnešních ploutvenek u fosilních zástupců. Archaické typy se totiž mohly vyznačovat značně odlišnou stavbou vnitřních orgánů a jejich vymřením se zcela ztratila informace o počátečních vývojových fázích celého kmene. V budoucnu se může ukázat, že takovou formou je i střednokambrický druh Amiskwia sagittiformis. Blízká dnešním ploutvenkám se jeví již spodnoordovická Titerina rokycanensis, popřípadě svrchnokambrický Prooneotodus? tenuis. Je tedy vysoce pravděpodobné, že ploutvenky představují velmi starou skupinu, která se objevuje již počátkem paleozoika. Vzhledem k vzácné fosilizaci nelze ovšem vyloučit ani jejich prekambrický původ. Představám o značné starobylosti skupiny odpovídá také její izolované postavení mezi dnešními živočišnými kmeny. 3)
Literatura
Bone Q., Kapp H., Pierrot-Bults A. C. (eds.): The biology of Chaetognaths, 1–173. Oxford University Press, Oxford 1991Casanova J. P., Duvert M., Goto T.: Emergence of limb-like appendages from fins in chaetognaths. C. R. Acad. Sci. Paris, Life Sciences 318, 1167–1172, 1995
Duvert M., Salat C.: Ultrastructural studies of the visceral muscles of Chaetognaths. Acta Zoologica (Stockholm) 76, 75–87, 1995
Halanych K. M.: Testing hypotheses of chaetognath origins: long branches revealed by 18S ribosomal DNA. Syst. Biol. 45, 223–246, 1996
Hyman L. H.: The enterocoelous coelomates – phylum Chaetognatha. The invertebrates: smaller coelomate groups 5, 1–71. McGraw-Hill, New York 1959
Kraft P., Mergl M.: Worm-like fossils (Palaeoscolecida; ?Chaetognatha) from the Lower Ordovician of Bohemia. Sbor. geol. věd, Paleont. 30, 9–36, 1989
Szaniawski H.: Chaetognath grasping spines recognized among Cambrian protoconodonts. J. Paleont. (Tulsa) 56, 806–810, 1982
Telford M. J., Holland W. H.: The phylogenetic affinities of the Chaetoghaths: a molecular analysis. Mol. Biol. Evol. 10, 660–676, 1993
Terazaki M., Miller C. B.: Reproduction of meso- and bathypelagic chaetognaths in the genus Eukrohnia. Marine Biology 71, 193–196, 1982
Obrázky







Poznámky
SLOVNÍČEK
akrozom
Ke stažení
 Článek ve formátu PDF [1,79 MB]
Článek ve formátu PDF [1,79 MB]
O autorech
Petr Kraft
Jiří Král
Doporučujeme

Rostliny vyprávějí o lidech

O prázdnech v nás