Netušená síla slabých vazeb
 Zatímco o kovalentních vazbách v molekulách se na základní či střední škole učili všichni čtenáři Vesmíru, povědomí o nekovalentních interakcích a molekulárních klastrech je mnohem omezenější. Přitom neexistuje jediný významný proces probíhající v živém prostředí, který by nekovalentními interakcemi nebyl od počátku až do konce ovlivněn.
Zatímco o kovalentních vazbách v molekulách se na základní či střední škole učili všichni čtenáři Vesmíru, povědomí o nekovalentních interakcích a molekulárních klastrech je mnohem omezenější. Přitom neexistuje jediný významný proces probíhající v živém prostředí, který by nekovalentními interakcemi nebyl od počátku až do konce ovlivněn.
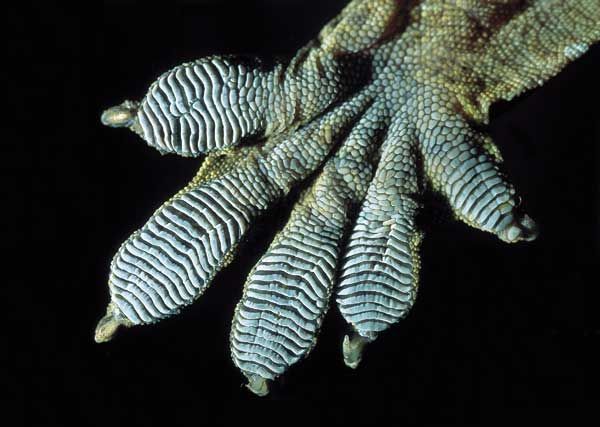
Nekovalentní interakce jsou mnohem slabší než interakce kovalentní. Nikoliv přesto, ale právě proto hrají tak významnou úlohu v procesech ovládajících živou hmotu. Jsou však stejně podstatné i v jiných oblastech přírodních věd, v chemii či fyzice, a dokonce i v živočišném světě. Překvapivá schopnost gekona udržet se na kolmých a převislých plochách je možná díky nekovalentním interakcím mezi keratinovými vlákny chodidel (obr. 3) a molekulami skla. Všude, kde se k sobě přiblíží dvě nebo více molekul, vznikají v důsledku nekovalentních interakcí molekulové klastry, které ovlivní chování, vlastnosti i reaktivitu systému.
Vše začíná u vody
 Většina chemických reakcí se odehrává v kapalné fázi, většinou ve vodném prostředí. Sama existence kapalného (a zvláště vodného) prostředí je důsledkem existence nekovalentních interakcí. Molekuly rozpouštědla se navzájem přitahují a to je nezbytnou podmínkou vzniku kapalné fáze. Za extrémně nízkých teplot a extrémně vysokých tlaků existují i vzácné plyny v kapalném stavu; voda je naopak kapalná za normálních teplot a tlaků.
Většina chemických reakcí se odehrává v kapalné fázi, většinou ve vodném prostředí. Sama existence kapalného (a zvláště vodného) prostředí je důsledkem existence nekovalentních interakcí. Molekuly rozpouštědla se navzájem přitahují a to je nezbytnou podmínkou vzniku kapalné fáze. Za extrémně nízkých teplot a extrémně vysokých tlaků existují i vzácné plyny v kapalném stavu; voda je naopak kapalná za normálních teplot a tlaků.
Voda má zásluhou nekovalentních interakcí velmi specifické vlastnosti (neobvykle vysoký bod varu a bod tání, vysokou dielektrickou konstantu, vysoké povrchové napětí…), které se dramaticky liší od vlastností jiných systémů se stejným počtem elektronů, například amoniaku, metanu či fluorovodíku. Očekávali bychom, že vlastnosti různých molekul se stejným počtem elektronů by měly být podobné. Opak je však pravdou. Jako příklad uveďme body varu: fluorovodík 20 °C, voda 100 °C, amoniak –33 °C a methan –161 °C.
Důvodem specifických vlastností vody je existence silných nekovalentních interakcí, vodíkových vazeb (viz dále), které se u ostatních látek nevyskytují, nebo jsou mnohem slabší. Spekuluje se, že právě neobvyklé vlastnosti vodného prostředí umožnily vznik života – stále si jen těžko představujeme jiné podmínky, v nichž by mohl život vzniknout.
Záruka flexibility DNA
 Na obr. 1 je znázorněna struktura DNA. Její dvoušroubovicové uspořádání je zásadně důležité pro schopnost DNA uchovávat a přenášet (kopírovat) dědičnou informaci: Po enzymy vyvolaném otevření DNA a rozpojení obou vláken dojde k syntéze chybějícího druhého vlákna obou dceřiných molekul, takže výsledkem jsou dvě identické kopie původní dvoušroubovice. Genetická informace daná pořadím bází nukleových kyselin na jednotlivém vláknu se tak zkopíruje na dvě dceřiná vlákna.
Na obr. 1 je znázorněna struktura DNA. Její dvoušroubovicové uspořádání je zásadně důležité pro schopnost DNA uchovávat a přenášet (kopírovat) dědičnou informaci: Po enzymy vyvolaném otevření DNA a rozpojení obou vláken dojde k syntéze chybějícího druhého vlákna obou dceřiných molekul, takže výsledkem jsou dvě identické kopie původní dvoušroubovice. Genetická informace daná pořadím bází nukleových kyselin na jednotlivém vláknu se tak zkopíruje na dvě dceřiná vlákna.
Struktura DNA je určena různými typy nekovalentních interakcí mezi stavebními bloky DNA, kterými jsou báze nukleových kyselin (guanin, cytosin, adenin a thymin), cukry a fosfátové skupiny, a také mezi těmito stavebními bloky a molekulami vody a protiionty (Na+, Mg2+...).
Přenos genetické interakce v DNA i v RNA je názorným příkladem, proč si příroda vybrala slabé nekovalentní interakce jako rozhodujícího hráče. Aby plnila svou funkci, nesmí být DNA či RNA rigidní, ale relativně snadno otvíratelná. Proces otevření a uzavření musí být reversibilní (probíhat oběma směry) a vysoce přesný. Nekovalentní interakce, které tyto procesy řídí, musí být na jedné straně dostatečně silné, aby zajistily preferenční vazbu guaninu s cytosinem a adeninu s thyminem, a na druhé straně musí být dostatečně slabé, aby mohlo dojít k otevření dvoušroubovice. Je to jako v pohádce o chytré horákyni. Ta také musela splnit zdánlivě protichůdné podmínky: Přijít ani ve dne, ani v noci, ani ustrojená, ani nahá, ani pěšky, ani na voze.
Stejné nekovalentní interakce se uplatní i v procesu molekulárního rozpoznání, kdy jedna molekula (např. guanin) zcela bezchybně a na velkou vzdálenost rozpozná komplementárního partnera (tedy cytosin). Přesnost tohoto procesu při replikaci musí být neuvěřitelně vysoká, toleruje se jen jedna chyba z milionu, a ta se obvykle opraví za účasti opravných enzymů, které pracují (jak jinak) s nekovalentním rozpoznáním chyby.
Uvědomíme-li si, že větší výskyt chyb může vést až ke karcinogennímu procesu, pochopíme zcela zásadní úlohu, kterou nekovalentní interakce v biodisciplínách hrají. Tyto interakce určují i strukturu dalších biomakromolekul včetně bílkovin, a také komplexů bílkovin s DNA a s ligandy (léky, ale také např. karcinogenními látkami).
Odkud se berou?
Kovalentní vazba vznikne, dojde-li k překryvu elektronových oblaků dvou systémů s nespárovanými elektrony. Interakcí dvou atomů vodíku dojde ke vzniku molekuly vodíku, interakcí dvou metylových radikálů vznikne molekula etanu... Energie kovalentních vazeb se obvykle pohybuje mezi 40 a 100 kcal/mol. Při tvorbě kovalentní vazby se toto množství energie uvolní; a naopak, chceme-li kovalentní vazbu rozbít, musíme takové množství energie dodat. Podmínkou vzniku kovalentní vazby je dostatečné přiblížení obou systémů (na méně než 2 Å2)).
Ze zkušenosti však víme, že nekovalentní interakce se projevují i na mnohem větší vzdálenosti mezi systémy (až desítky Å). Je tedy zřejmé, že elektronový překryv, který s rostoucí vzdáleností velmi rychle mizí, nemůže být podstatou nekovalentních interakcí. Analyzujeme-li všechny možné zdroje přitahování, zjistíme, že jediným původcem nekovalentních interakcí mohou být elektrické vlastnosti molekul.
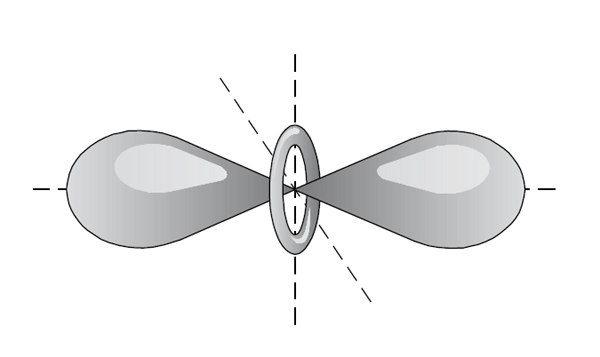
 Nerovnoměrné rozložení elektronů v molekule s různými atomy vede ke vzniku permanentního elektrického momentu, většinou dipólového. U molekul s vyšší symetrií se může uplatnit quadrupólový (případně vyšší multipólový) moment. Permanentní elektrické momenty na sebe působí (přitažlivě nebo odpudivě podle geometrického uspořádání obou systémů) a výsledná energetická složka se nazývá coulombická nebo elektrostatická interakční energie. Interakční energie dvou polárních systémů s dipólovými momenty je úměrná součinu obou momentů a nepřímo úměrná třetí mocnině vzdálenosti mezi molekulami.
Nerovnoměrné rozložení elektronů v molekule s různými atomy vede ke vzniku permanentního elektrického momentu, většinou dipólového. U molekul s vyšší symetrií se může uplatnit quadrupólový (případně vyšší multipólový) moment. Permanentní elektrické momenty na sebe působí (přitažlivě nebo odpudivě podle geometrického uspořádání obou systémů) a výsledná energetická složka se nazývá coulombická nebo elektrostatická interakční energie. Interakční energie dvou polárních systémů s dipólovými momenty je úměrná součinu obou momentů a nepřímo úměrná třetí mocnině vzdálenosti mezi molekulami.
Přitahují se také polární systémy (s permanentními elektrickými momenty) s nepolárními systémy, např. atomy vzácných plynů. V důsledku vzájemného působení dojde v nepolárním systému ke vzniku indukovaného momentu vyvolaného elektrickým polem polární molekuly a mezi permanentním a indukovaným momentem dojde k interakci. Tato indukční interakce je méně významná než interakce coulombická.
Zkapalnění vzácných plynů ukázalo, že musí existovat přitažlivá interakce i mezi nepolárními systémy, které nemají žádný permanentní elektrický moment. Nazývá se Londonova disperzní interakce a působí mezi všemi systémy. Byla vysvětlena až pomocí kvantové mechaniky a lze ji zjednodušeně interpretovat následovně: V důsledku oscilací atomových jader a elektronového obalu dojde ke vzniku časově proměnného dipólového momentu, který následně indukuje moment i ve druhém systému. Mluvíme tak o interakci časově proměnného a indukovaného momentu a výsledná disperzní interakce je, stejně jako indukční interakce, vždy přitažlivá.
Londonova disperzní interakce se někdy zaměňuje s Van der Waalsovými (vdW) interakcemi, které jsou široké veřejnosti známější a učí se o nich i na střední škole. Dochází tady k částečnému matení pojmů. Na konci minulého století byly vdW interakce synonymem nekovalentních interakcí; v současnosti preferujeme název nekovalentní interakce. Název vdW interakce se správně užívá pro součet přitažlivých disperzních a odpudivých výměně-repulsních interakcí (viz dále).
Všechny uvedené složky mohou být při určitém geometrickém uspořádání přitažlivé, a tak musí existovat nějaká protiváha – energie, která zabrání, aby se systémy mohly příliš přiblížit a prostorově splynout. Tato energie, nazývaná výměně-repulsní, je stejně jako energie disperzní kvantové povahy a její původ je analogický vzniku kovalentní vazby.
V případě kovalentní vazby ale reagují systémy s neúplně zaplněnou elektronovou sférou (nespárovanými elektrony), a tak dojde ke zvýšení elektronové hustoty mezi atomy (ve vazebném orbitalu) a vznikne vazba. V případě výměně-repulsní energie interagují systémy se zaplněnou elektronovou sférou, elektrony nemohou obsadit již obsazený vazebný orbital a obsazují antivazebný orbital. V důsledku toho dojde ke snížení elektronové hustoty mezi atomy a oslabení vazebnosti, což vede k odpuzování systémů.
Vodíkové vazby a nepravé vodíkové vazby
Vodíkové vazby patří mezi nejsilnější typy nekovalentních interakcí. Ve vodíkové vazbě X-H...Y je vodík sdílen (vytváří jakýsi můstek) mezi elektronegativními atomy X a Y, přičemž atom Y (nebo skupina atomů) obsahuje přebytek elektronů. Atom Y je protonový akceptor. Jde většinou o atom s volným elektronovým párem (O, N, F,...), ale může to být také systém s elektrony v delokalizovaných (aromatických) orbitalech (benzen). Charakteristickým strukturním rysem je prakticky lineární uspořádání atomů X-H-Y.
Kovalentní vazba X-H je polarizovaná (atom X je elektronegativnější než vodík) a to značí, že vodík nese částečný kladný náboj. Atom Y vzhledem k přítomnosti volných elektronových párů nese naopak částečný záporný náboj a přitažlivá elektrostatická interakce mezi těmito náboji představuje převážnou část interakce vodíkové vazby. Komplexy s vodíkovou vazbou patří mezi nejběžnější a nejsilnější (5–10 kcal/mol) nekovalentní komplexy, přesto je vodíková vazba stále asi desetkrát slabší než kovalentní vazby (viz výše).
Vznik vodíkové vazby X-H...Y nejlépe vysvětluje tzv. přenosový (anglicky „charge- transfer“) model, podle kterého dochází k přenosu elektronové hustoty z elektronových párů protonového akceptoru (Y) do antivazebného orbitalu vazby X-H protonového donoru. (Vazebný orbital X-H je nepoužitelný, protože je plně obsazen elektrony.) Zvýšení elektronové hustoty v antivazebném orbitalu vede k oslabení vazby X-H, jejímu prodloužení a následně ke zmenšení vazebné vibrace X-H (červený posun).
Červený posun vazebné vibrace byl dlouho považován za charakteristiku jednoznačně označující vznik vodíkové vazby. V naší laboratoři jsme však ukázali3), že vznik vodíkové vazby může být provázen také nárůstem vazebné vibrace X-H (modrý posun). Tzv. nepravá vodíková vazba byla pokusně detekována nebo teoreticky předpovězena jak v plynné, tak i v kapalné a pevné fázi.
Vlastnosti komplexů se standardní (červenou) nebo nepravou vodíkovou vazbou jsou velmi podobné, pokud jde o strukturu, geometrii, stabilizační energii i absolutní hodnotu velikosti posunu vibrační frekvence X-H. Objevení nepravé vodíkové vazby zkomplikovalo definici vodíkové vazby. IUPAC ji proto změnil tak, že připouští posun vibrační frekvence X-H jak k nižším (červený posun), tak k vyšším hodnotám (modrý posun).
Vodíková vazba, ať už s červeným, nebo modrým posunem, není jediným příkladem nekovalentní vazby, na které se podílí atom vodíku. Velmi překvapivá interakce typu XH... H-Y byla nalezena v metalických komplexech a později potvrzena i v heteroboranových komplexech. Vysvětlení této na první pohled nepochopitelné anomálie je prosté: Je-li vodík vázán na elektronegativní atom, nese částečný kladný náboj. Je-li však vodík kovalentně vázán na elektropozitivní prvek (bor, alkalické kovy, tranzitní kovy), nese naopak částečný záporný náboj. Vazba X-H...H-Y, kde X a Y jsou elektronegativní a elektropozitivní elementy, je tak způsobena elektrostatickým přitahováním vodíkových atomů nesoucích opačné náboje. Tato tzv. dvouvodíková vazba (angl. dihydrogen bond) může dosahovat až síly standardní vodíkové vazby (~ 5 kcal/mol).
Disperzní interakce v nukleových kyselinách a struktura DNA
Disperzní interakce (viz výše) je vždy přitažlivá. Většinou je však spojována se stabilizací dimerů vzácných plynů, kde představuje jedinou přitažlivou složku. Vzhledem k tomu, že stabilizační energie dimerů vzácných plynů se pohybuje od 0,1 do zhruba 1 kcal/mol, soudilo se, že disperzní energie nehraje u velkých systémů významnou úlohu.
Až přesné výpočty interakčních energií rozsáhlých systémů (páry bází nukleových kyselin, páry aminokyselin) ukázaly, že stabilizační energie patrových komplexů těchto systémů (dvou bází nad sebou) je srovnatelná se stabilizační energií planárních komplexů s násobnými vodíkovými vazbami. Např. stabilizační energie patrových komplexů guaninu s cytosinem a adeninu s thyminem (18 a 13 kcal/mol) v DNA je zhruba srovnatelná se stabilizační energií planárních komplexů (29 a 16 kcal/mol) stejných systémů majících tři (guanin-cytosin), respektive dvě (adenin-thymin) silné vodíkové vazby.
Tento závěr je překvapivý a má významné důsledky. Ten nejdůležitější se týká původu stabilizace molekuly DNA i bílkovin. V literatuře stále převládá názor, že stabilita DNA je dána především silnými vodíkovými vazbami v párech guanin-cytosin a adenin-thymin. Ukazuje se však, že tyto stabilizační interakce (mající původ v elektrostatických interakcích) jsou kompenzovány hydratačními energiemi, a stabilita DNA je tak určena především patrovými interakcemi mezi bázemi nukleových kyselin. Ukázali jsme, že patrové interakce jsou především určeny disperzními interakcemi a ty (na rozdíl od vodíkových vazeb) jsou hydratačními energiemi ovlivněny daleko méně. Velká stabilita patrových párů také vysvětluje překvapivou stabilitu modifikovaných molekul DNA, ve kterých je kanonický pár silně polárních bází nukleových kyselin nahrazen párem nepolárních bází.
Struktura bílkovin
Některé bílkoviny obsahují jádro tvořené hydrofobními aminokyselinami („nemají rády“ vodu; jsou málo polární a ve vodě těžko rozpustné). V učebnicích biochemie nalezneme definici hydrofobního jádra jako důsledek hydrofobních interakcí molekul vody obklopujících bílkovinu. Tato stabilizace je tak entropické (vycházející z rozdílné uspořádanosti), a nikoliv enthalpické (vycházející z rozdílné energie interakce) povahy. Interakce aminokyselin v hydrofobním jádru se pak očekává velmi malá, nebo dokonce repulsní povahy.
Přesné výpočty interakčních energií aminokyselin v hydrofobním jádře bílkoviny rubredoxinu však ukázaly, že všechny párové interakční energie jsou přitažlivé a stabilizační energie párů aromatických aminokyselin dosahují hodnot 5–8 kcal/mol. Vodíková vazba v dimeru vody přitom nepřesahuje hodnotu 5 kcal/mol. Toto zjištění samozřejmě mění náš názor na charakter interakcí v hydrofobním jádru proteinu. Je totiž zřejmé, že aminokyseliny jsou uspořádány na energetickém základě (jednotlivé páry zaujímají téměř optimální strukturu, jakou by měly v plynné fázi). Tento závěr je významný proto, že hydrofobní jádro patrně hraje důležitou úlohu při životně důležitém procesu sbalování („folding“) bílkovin.
Nekovalentní interakce nepůsobí jen mezi molekulami, ale i v rámci jednotlivých molekul. V malých rigidních systémech je jejich úloha samozřejmě omezená, ale u velkých flexibilních systémů, např. peptidů či bílkovin, se jak vodíkové vazby, tak disperzní interakce významně uplatňují a spoluurčují jejich strukturu.
Interakce s cizorodými látkami
Nejdůležitějšími cizorodými látkami jsou bezesporu léky. Ve velké většině případů jsou cílem léků bílkoviny (hlavně enzymy), v některých případech také DNA či RNA. Aktivita léků je způsobena tzv. kompetitivní inhibicí. Lék se váže reversibilně do aktivního místa, a brání tak navázání původního substrátu. Zřejmě prvním požadavkem je, aby lék měl vhodný tvar, který napodobuje tvar aktivního místa. Druhým požadavkem je, aby se lék do aktivního místa navázal.
Velká většina léků se váže do aktivního místa nekovalentně. Na obr. 2 je znázorněn komplex HIV proteázy s inhibitorem (atazanavir). Z obrázku je zřejmé, že inhibitor velmi dobře vyplňuje aktivní místo bílkoviny, a také to, že inhibitor je vázán nekovalentně. Velmi zjednodušeně lze říci, že optimalizaci účinku léku lze převést na maximalizaci změny volné energie tvorby komplexu lék – biologicky významná látka (protein, nukleová kyselina, membrána…). Čím je změna vazebné volné energie větší, tím bude inhibitor (lék) účinnější.
Tento proces je velmi komplexní a kromě samotné nekovalentní vazby léku s biologickou látkou hrají významnou úlohu ještě hydratace/dehydratace léku i látky a deformace obou systémů. Nekovalentní vazba léku s aktivním místem však představuje klíčový proces a pochopení podstaty této vazebnosti je zásadní pro racionální návrh nových léků.
Poznámky
1) P. Hobza, K. Müller-Dethlefs, Non-Covalent Interactions – Theory and Experiment, RSC Publishing; Cambridge, 2009
2) Å, angström (10–10 m), je vhodná jednotka vzdálenosti pro měření v molekulách.
3) P. Hobza, Z.Havlas: Chem. Rev. 100, 4253, 2000
Molekulární klastry
Chemie zkoumá strukturu, vlastnosti a reaktivitu molekul. Při chemické reakci se vazby v molekulách přesunují a vytvářejí nové molekuly s vlastnostmi zcela odlišnými od vlastností původních molekul. Molekuly však spolu mohou reagovat i bez vzniku a zániku kovalentních vazeb. Produktem takové reakce je molekulární shluk (klastr).
Molekuly jsou v klastru vázány pohromadě mnohem slabšími nekovalentními interakcemi a vlastnosti molekul vytvářejících komplex se jen málo liší od vlastností původní molekuly. Počet druhů molekul je obrovský (uvádí se několik miliard), avšak počet molekulových klastrů je prakticky neomezený, protože prakticky každá molekula může intereagovat s jinou za vytvoření klastru. Molekulové klastry mohou obsahovat dva nebo také více systémů jednoho nebo více druhů a celkový počet molekul může být obrovský. V tom případě dochází za určitých teplot a tlaků ke vzniku kondenzované fáze (např. molekulových krystalů).
Ve většině případů jsou vlastnosti interagujících systémů v klastru zachovány. Za určitých podmínek však vede vznik molekulového klastru ke změně vlastností interagujících systémů. Jako příklad uveďme komplex dioxanu s trinitrometanem. Jedovatá látka dioxan interaguje s dvěma molekulami výbušné látky trinitromethanem, vytváří dvě velmi silné vodíkové vazby typu C-H…O. Nově vzniklý klastr se uspořádá do molekulového krystalu a překvapivě není ani toxický, ani výbušný!
Ke stažení
 článek ve formátu pdf [455,8 kB]
článek ve formátu pdf [455,8 kB]
O autorech
Pavel Hobza
Zdeněk Havlas
Doporučujeme

Rostliny vyprávějí o lidech

O prázdnech v nás