













Život bez hlavy
| 6. 1. 2016Hlava je pro nás obratlovce bezesporu zásadní částí těla, která ale nemusí být vlastní mnohým jiným skupinám živočichů. Jedním z nich je i kopinatec, jenž vypadá jako bezhlavá rybka a obratlovcům se podobá svým embryonálním vývojem a vnitřní stavbou těla. I přes zjevnou nepřítomnost hlavy lze u kopinatce vysledovat znaky vypovídající o tom, jakými směry se evoluce této části těla ubírala.
Naše hlava je složitou strukturou, kterou lze charakterizovat přítomností smyslových orgánů, mozku, ústí dýchací soustavy, úst obsahujících aparát pro příjem a zpracování potravy; a to vše je navíc kryté kostěnou schránkou. Podobně jako my, existují živočichové (např. ploštěnci, měkkýši nebo členovci), kteří mají hlavu také dobře definovanou, i když jinak a specificky pro danou skupinu. Dále existuje celá plejáda živočišných skupin, které se bez hlavy docela dobře obejdou (například někteří mořští živočichové – mořské houby, medúzy, korály, hvězdice či mechovky) a pak zde máme živočichy takříkajíc napůl cesty, tzn. jejich tělo sice obsahuje přední a zadní konec, ale jasně definovanou hlavu nemají. Jedním z takovýchto živočichů je kopinatec.
Kopinatci s obratlovci sdílí mnohé znaky. Především je to struna hřbetní, tzv. chorda, která slouží jako výztuha těla, trubicovitá nervová soustavu umístěná na zádové straně těla, dále jasně diferencovaná ocasní část sloužící k pohybu a tělo tvořené segmentovanou svalovinou. Ze znaků přítomných v přední části je to hltan, coby jasně odlišitelný oddíl trávicí trubice, nebo endostyl, struktura obdobná štítné žláze obratlovců. Kromě těchto znaků sdílí kopinatci s obratlovci také raný embryonální vývoj, který je pro pochopení evoluce obratlovců naprosto zásadní. Lze z embryonálního vývoje kopinatce vyvodit, jaké důležité embryonální struktury pomáhaly utvářet hlavu obratlovců?
Mozek a oči
Kopinatec stráví většinu svého dospělého života zahrabaný v písku. Skryt před zraky potenciálních predátorů, nepotřebuje pro své přežití složitě vyhodnocovat únikové strategie. Jeho nervová soustava a vizuální aparát tak nebyly v evoluci pod selekčním tlakem „musím vidět predátora dříve, než on uvidí mě“ a případně „musím nalézt nejvhodnější únikovou cestu“, jako tomu bylo u jiných živočichů, kteří se během života pohybují ve volném prostoru.
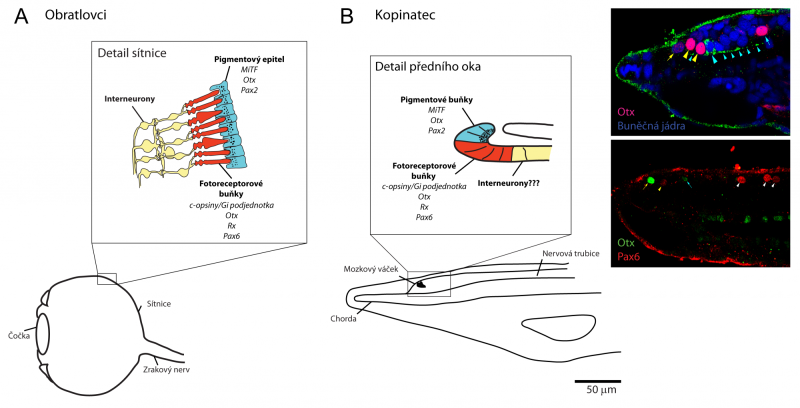
Přední zesílená část nervové trubice kopinatce, tzv. mozkový váček. Na rozdíl od mozku obratlovců, tvořeným miliony až miliardami neuronů, čítá mozkový váček kopinatce pouze stovky neuronů z celkového počtu asi dvaceti tisíc neuronů v celé nervové soustavě. Z morfologického hlediska tak není u kopinatce jednoduché rozlišit, kde končí „mozek“ a začíná jeho „mícha“.
Při výzkumu nervové soustavy kopinatce byla v historii (přelom 19. a 20. století) větší pozornost věnována právě studiu míchy, mimo jiné i proto, že je schopna částečně fungovat nezávisle na mozkovém váčku (vykazuje například reakci na světelné stimuly, vedoucí ke snaze skrýt se do písečného podloží). Mozkový váček tak zůstával mimo zájem badatelů. Přelomovými pracemi byly studie Thurstona Lacalliho, který provedl několik anatomických rekonstrukcí vyvíjejícího se mozkového váčku kopinatce. Ve svých pracích integroval elektronmikroskopická pozorování tisíců po sobě jdoucích řezů mozkových váčků larev kopinatců. Lacalli a kol. v mozkovém váčku kopinatce identifikovali struktury, které označili za srovnatelné s obratlovčími mozkovými strukturami (např. přední oko kopinatce/oči obratlovců; lamelární orgán kopinatce/epifýza obratlovců; infundibulární orgán kopinatce/neurohypofýza obratlovců; přilehlé dorzální a ventrální shluky neurosekrečních buněk kopinatců/hypothalamo-hypofyzární systém obratlovců). Všechny tyto závěry byly učiněny pouze na základě morfologie neuronů a postupně jsou doplňovány probíhajícími studiemi sledujícími expresi neurotransmiterů, které definují různé populace neuronů.
Kromě anatomických pozorování paralelně probíhaly i studie exprese genů. Ve vyvíjejícím se mozkovém váčku kopinatce byla pozorována exprese genů, které jsou důležité pro vývoj a ustavení jednotlivých částí mozku obratlovců. Propojení těchto genů do logiky genových regulačních sítí však není mezi kopinatcem a obratlovci shodné. Tyto rozdíly jsou považovány za důležité pro evoluci mozku obratlovců a jsou podrobeny dalšímu zkoumání.
Asi nejlépe se podařilo ověřit shodu mezi předním okem kopinatce a komorovým okem obratlovců. Kopinatec má v mozkovém váčku na rozdíl od obratlovců oko pouze jedno a jeho morfologie je velmi jednoduchá – skládá se z asi deseti fotoreceptorových a asi pěti přilehlých pigmentových buněk a nelze u něj najít strukturu podobnou čočce. Jeho zobrazovací schopnosti jsou velmi nízké, pravděpodobně poskytuje pouze informaci o přítomnosti nebo absenci světla, čili o tom, zda se neblíží stín, signalizující predátora.
Na vývoji a funkci předního oka kopinatce a komorových očí obratlovců se však podílí stejné geny, proto se pravděpodobně jedná o evolučně velmi konzervovaný program.
Ve zkratce se tedy dá říct, že z hlediska mozku a vizuálního aparátu lze najít mezi kopinatcem a obratlovci mnoho shodných znaků, které byly pravděpodobně zděděny od společného předka. Rozdíly pak jsou nejspíš způsobeny ztrátou některých funkcí centrální nervové soustavy u kopinatce, které nebyly vzhledem k jeho způsobu života potřebné a hlavně pak získanými vylepšeními u předků v linii vedoucí k dnešním obratlovcům.
ČTĚTE TAKÉ: Stále záhadný kopinatec a Čtvrtá vrstva
Neurální lišta a evoluce lebky
Neurální lišta je populace buněk, která vzniká při utváření nervové trubice u embryí obratlovců (obr. 2). Tato populace buněk posléze putuje celým zárodkem a dává vznik mnoha tkáním a buněčným typům. V hlavě neurální lišta kromě jiného tvoří chrupavky, lebeční kosti, zuby (kromě skloviny), pojivové tkáně a škáru. Neurální lišta je tedy zásadní embryonální strukturou, neboť hlavu jako takovou utváří.
Kopinatci nemají lebku ani neurální lištu. Studiem embryonálního vývoje kopinatce se zjistilo, že faktory zodpovědné za specifikaci a migraci buněk neurální lišty obratlovců se u kopinatce sice nacházejí, ale netvoří správné vzájemné interakce. O to více je překvapivé, že aparát kolem úst tvoří u kopinatců buněčná chrupavka, tj. tkáň, která je u obratlovců derivátem právě buněk neurální lišty a která je základním materiálem pro stavbu čelistí, jazylky či ušních kůstek. Naproti tomu chrupavky ústního aparátu jsou u kopinatců tvořeny buňkami mezodermu, tedy buněčné populace, ze které vznikají mimo jiné i svaly.
Předpokládáme-li, že absence neurální lišty a tvorba chrupavek ústního aparátu z mezodermu reprezentují u kopinatce do jisté míry původní stav znaku, je nasnadě uvažovat, že evoluce lebky obratlovců na svém počátku vyžadovala dva důležité momenty. Prvním je nabytí potenciálu pro tvorbu chrupavky z neurální lišty a druhým rozšíření tohoto potenciálu i do jiných míst budoucí lebky, přičemž první moment nemusí nutně předcházet tomu druhému.
Kde se ale vzala neurální lišta? Nedávný výzkum pláštěnce sumky trubičkovité poukázal na přítomnost buněk, které se svými vlastnostmi podobají neurální liště obratlovců. Uzavírající se nervová trubice sumky dává vznik několika buňkám, ze kterých vznikají periferní nervy ne nepodobné spinálním gangliím obratlovců. Z pozice pláštěnců, coby sesterské linie obratlovců, lze usuzovat, že předek obratlovců již tvořil buňky neurální lišty, které pravděpodobně měly potenciál generovat buněčnou chrupavku, a tato posléze vytvořila základ pro kostěnou lebku.
Evoluce smyslových orgánů
Do značné míry opomíjenými, zato ale velmi důležitými strukturami, které pomáhají spoluutvářet hlavu obratlovců, jsou hlavové plakody. Podobně jako neurální lišta vznikají ze společného základu (preplakodální oblast) během utváření nervové trubice a posléze migrují, interagují s okolními tkáněmi a diferencují do mnoha buněčných typů.

Dávají vznik smyslovým orgánům (jako např. vnitřnímu uchu, čichovým vakům, oční čočce či postranní čáře), gangliím hlavových nervů či přednímu laloku podvěsku mozkového (tzv. adenohypofýze). Díky své poloze a potenciálu tvořit množství tkání se předpokládá, že do určité míry sdílejí evoluční původ s neurální lištou.
U kopinatce k tvorbě plakod nedochází. O to více je zajímavé, že dvě části jeho těla lze do určité míry srovnat s plakodami obratlovců, jakými jsou adenohypofýza a čichové vaky. Jedná se o rostrální ektoderm a o preorální jamku. Rostrální ektoderm obsahuje množství vzájemně oddělených senzorických neuronů, které exprimují transkripční faktory důležité pro vývoj čichových vaků obratlovců a také nejméně jeden typ obratlovčího genu kódujícího čichový receptor. Preorální jamka zase obsahuje obrvené buňky, které exprimují faktory důležité pro vývoj adenohypofýzy obratlovců. Preorální jamku lze srovnávat s adenohypofýzou také na základě jejího pozdějšího spojení s mozkovým váčkem. Toto spojení silně připomíná vyvíjející se podvěsek mozkový obratlovců. Co je ovšem zvláštní, preorální jamka vzniká z výchlipky přední části střeva, tedy z naprosto odlišné buněčné populace, než z jaké bychom její vznik bývali očekávali .
Společný předek kopinatců a obratlovců tedy zřejmě neměl definovanou preplakodální oblast, ale nesl neurosekretorické a senzorické neurony koncentrované do jasně definovaných orgánů. Důležitým milníkem v evoluci obratlovců přitom bylo spuštění exprese transkripčních faktorů, které se staly zodpovědnými za vývoj preplakodální oblasti a za diferenciaci této oblasti do jednotlivých plakod.
Tělní asymetrie a evoluce úst
Oproti výše popsaným znakům, které jsou u kopinatců v primitivním či zjednodušeném stavu a u obratlovců ve stavu odvozeném, představuje tělní asymetrie kopinatců určité specifikum. Nehovořili bychom zde o ní, kdyby zásadním způsobem nezasahovala do vývoje jeho přední části těla. U obratlovců se tělní asymetrie projevuje nejvíce v pozici a anatomii vnitřních orgánů (žaludek a slezina nalevo, játra napravo, plíce a srdce asymetricky utvářené). Vnější stavba těla je však většinou symetrická (výjimkou může být např. platýz). Naproti tomu jsou larvy kopinatců jedněmi z nejvíce asymetrických živočichů vůbec, a to jak svou vnější tak vnitřní stavbou. Asymetrické je v podstatě celé tělo. Svaly na levé a pravé straně, stejně jako nervy, které do nich vstupují, jsou navzájem posunuty. Tato nesouměrnost je ale nejvíce viditelná v přední části, kde ústa a preorální jamka vznikají nalevo, zatímco endostyl a kyjovitá (slizotvorná) žláza napravo. Všechny žaberní štěrbiny se zakládají napravo, a to tím způsobem, že se nejprve založí budoucí levé štěrbiny, které se přesunou zprava doleva a až poté se na pravé straně založí budoucí pravé štěrbiny. Během metamorfózy dojde k radikálnímu přesunu všech orgánů a k částečnému ustanovení bilaterální symetrie.
Ústa a ústní aparát hrají ústřední roli v evoluci strunatců a obratlovců. O to více je zarážející a matoucí situace u kopinatce, u něhož se ústa otevírají na levé straně. I přes svou levostrannost jsou nicméně srovnávána s ústy obratlovců a jejich neobvyklá poloha se vysvětluje přesunem původně uprostřed ležících úst doleva. Takovýto přesun ale předpokládá složité přesuny i dalších orgánů přidružených k ústům. Existují proto názory, že ústa v rámci strunatců nejsou srovnatelná, tedy že ústní otvory kopinatců a obratlovců nesdílejí společnou evoluční historii a kopinatčí ústa vznikla nezávisle na ústech obratlovců.
Podporu pro tento argument můžeme hledat v experimentu, při kterém porušíme funkci signální dráhy zodpovědné za levostranné umístění orgánů a levostrannou morfogenezi. Pokud by platilo, že ústa kopinatce se přesouvají nalevo z původní střední pozice, pak by takovýto experiment zabránil migraci úst doleva a vedl by k jejich tvorbě uprostřed. Ve skutečnosti ale vede ke ztrátě levostranných úst a preorální jamky a namísto nich vzniknou orgány, které se normálně vyvíjejí napravo – endostyl a kyjovitá žláza.
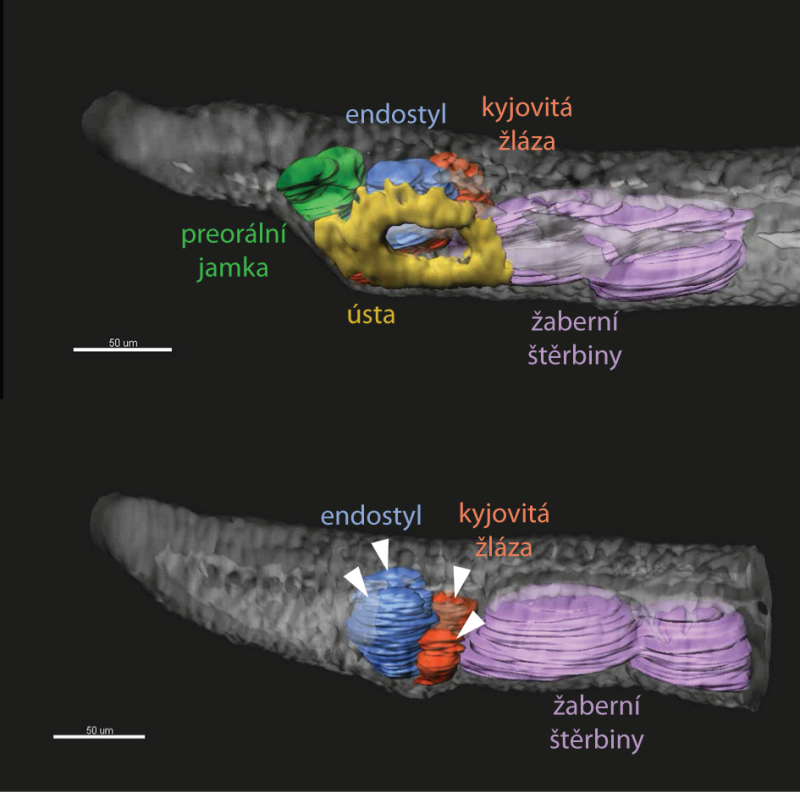
Celé tělo tedy vykazuje pravý izomerismus, tedy zrcadlový obraz pravé strany. Ústa a ostatní orgány přední části těla se během vývoje nikam nepřesouvají, ale primárně se zakládají asymetricky, nemusí tedy být nutně srovnatelná s ústy obratlovců.
Extrémní levopravá asymetrie je nicméně specifická pro kopinatce. Signální dráha zodpovědná za levostrannou morfogenezi je u kopinatců aktivní v mnohem větší míře a postihuje více orgánů než u obratlovců. Proč ale v evoluci kopinatců došlo k vývoji přes silně asymetrickou larvu, je otázkou, která zůstává nezodpovězená.
Evoluční nadstavba obratlovců – nová hlava
Hlava obratlovců je novotvarem, na jejíž stavbě se podílí mnoho odlišných buněčných populací. Zásadní pro evoluci hlavy se přitom stala oblast v přední části embrya spojená s uzavírající se nervovou trubicí, její diferenciace v hlavové plakody a neurální lištu a integrace těchto buněčných populací do složitějších celků.
U předka obratlovců dochází ke spojení plakod a mozku – adenohypofýzární plakoda a neurohypofýza dávají vznik podvěsku mozkovému, čočková plakoda a sítnice oku, čichové a sluchové vaky se sdružují s přilehlými částmi mozku. Plakody interagují s buňkami neurální lišty a spoluvytvářejí ganglia a hlavové nervy. Neurální lišta přitom utváří skeletální ochranu a podporu smyslovým orgánům, mozku a trávicí a dýchací trubici.
Celkem vzato se dá říci, že hlava obratlovců je výsledkem evoluce nových struktur vzniklých z nových buněčných populací plakod a neurální lišty. Všechny tyto interakce vedoucí k sestavení hlavy historicky souvisejí s přechodem předka obratlovců z živočicha pasivně filtrujícího potravu z okolního prostředí v živočicha s aktivním predátorským způsobem života. Tato změna způsobu života vedla k potřebě zdokonalit smyslové vnímání a postavit a modifikovat ústní aparát pro zachycení a příjem velké potravy.
Je důležité, že určité znaky této nové hlavy obratlovců lze vysledovat také u kopinatců. Kopinatci sice nemají specializované buněčné populace, jakými jsou plakody či neurální lišta, jejich tělní stavba ale vykazuje struktury, které jsou do jisté míry srovnatelné s obratlovčími strukturami tvořenými právě těmito buněčnými populacemi. A i když kopinatci žijí svůj život bez hlavy, stávají se důležitým zdrojem informací pro pochopení evoluce hlavy nás obratlovců.
Vopálenský P., Pergner J. a kol. (2012) Molecular analysis of the amphioxus frontal eye unravels the evolutionary origin of the retina and pigment cells of the vertebrate eye. Proc Nat Acad Sci USA 109, 15383-15388.
Soukup V. a kol. (2015) The Nodal signaling pathway controls left-right asymmetric development in amphioxus. EvoDevo 6, 5.
Černý R. (2010) Čtvrtá vrstva. Vesmír 89, 478-481.
Jandzik D. a kol. (2015) Evolution of the new vertebrate head by co-option of an ancient chordate skeletal tissue. Nature 518, 534-537.
Schlosser G. (2015) Vertebrate cranial placodes as evolutionary innovations – the ancestor’s tale. Curr Topics Dev Biol 111, 235-300.
Gans C. & Northcutt R.G. (1983) Neural crest and the origin of vertebrates: a new head. Science 220, 268-274.
O autorech
Jiří Pergner
Vladimír Soukup
Další články k tématu
Transplantace tváře. A co přijde pak?
Kolik váží lidská duše
Má hlava je včelín
Vědění - brzda radosti?
Vášeň a železná panna
Hlava – to nejcennější, co máme
O čem hovoří barevné hlavy želv
Želví hlava – enigma evoluční biologie
Hlas předpoví nemoc
Inteligentní bydlení pro schizofreniky
Eye tracking
Epileptické záchvaty a věštění z křišťálové koule
Doporučujeme

Když bahno teče jako ledovec

Příliš otevřené dveře
