










Epimutace proteinů
| 2. 2. 2015Chemické modifikace proteinů – záměna jedné aminokyseliny jinou – vedou ke změně jejich konformace a tudíž i aktivity. Náhodné mutace jsou většinou škodlivé a nevratné, zatímco epimutace – vlivem derivatizace aminokyselinových jednotek v hotovém proteinu – jsou cílené a dovolují velmi jemné nastavení aktivity proteinu dle potřeby. Vedle základní sady dvaceti aminokyselin mohou proteiny díky těmto možnostem obsahovat desítky aminokyselin dalších, a navíc lze tento repertoár dle potřeby za života proteinu i měnit.
Mějme řetězec aminokyselin – část proteinu sestávajícího z řetězce o několika stovkách aminokyselin:

(Řetězec je souvislý, mezery jsou jen pro snadnější orientaci. Písmena jsou jednopísmenové kódy aminokyselin, např. A=alanin, R=arginin apod.)
Záměna kterékoli aminokyseliny jinou, mutace, například K za R v poloze 14 může mít katastrofální následky pro strukturu a tím i funkci celého proteinu – a změna se často projeví i na celkové podobě organismu:

Zdůrazňujeme ono „může“ – některé aminokyseliny jsou zaměnitelné a nemusí se stát vůbec nic, anebo jen za určitých okolností či jen v některých typech buněk. Některé záměny jsou však přímo katastrofické – vzpomeňme u člověka srpkovou anémii nebo cystickou fibrózu. Takové změny jsou výsledkem náhodných a „nechtěných“ mutací DNA v tzv. kódující sekvenci genu pro příslušný protein, a svým nositelům připraví nepříliš snadný život.
K záměně aminokyselin však může dojít i posttranslačně, chemickou modifikací některé aminokyseliny v řetězci. Může se totiž stát, že úsek
bude v tomto složení jen v čerstvě syntetizované molekule proteinu. Jakmile je taková „panenská“ molekula „ošlehána životem“, tj. vsazena do kontextu ekosystému proteinů v buňce, dochází často k záměně některých aminokyselin za jiné a tím ke změně její konformace. Protože nejde o mutaci na úrovni DNA, nazývá se mutace tohoto druhu epimutací.
Některé příklady:

Na prvním řádku je opět nativní řetězec, na ostatních jsou hypotetické příklady epimutací – znázorněno řeckými písmeny: například ve 2. a 17. sloupci je aminokyselina arginin (R) pozměněna na dimetylarginin (ρ), nebo ve sloupci 16 je aminokyselina prolin (P) změněna na hydroxyprolin (π) anebo na izomer prolinu (ω). Další sloupce: 4. a 17. lysin na mono-, di, nebo trimetyllysin; 9. lysin navíc na acetyllysin; 10 serin na fosfoserin; 22. threonin na fosfothreonin.
Z 29 aminokyselin v daném řetězci jich takto může být teoreticky pozměněno nejméně 17 – pochopitelně v různých proteinech různě.
1. Je důležité si uvědomit, že chemická modifikace vytvoří zcela novou aminokyselinu: příklady na obr. 1 ukazují, že serin není fosfoserin a ani se mu svými vlastnostmi nepodobá, a podobně je to s různými deriváty lysinu. Zatímco však mutace způsobená změnami na DNA může způsobit nechtěné, znatelné či přímo zhoubné změny konformace proteinu, epimutace je cílená, ona musí způsobit takovou změnu konformace, v tom spočívá sám důvod změny. Ještě jednou: epimutace je mutace, ale přesně cílená na příslušné místo proteinu – jak, o tom níže. Navíc „běžné“ mutace mohou vybírat ze zásoby dvaceti aminokyselin (a ne vždy je taková „rošáda“ uskutečnitelná v jediném kroku), zatímco epimutace mohou vybírat z desítek možností – na aminokyseliny lze kromě výše uvedených skupin „připínat“ mastné kyseliny, geranyl, nejrůznější cukry, dokonce malé peptidy a proteiny apod. (viz Vojtěch Čermák, Šárka Motylová: Džungle epigenetických modifikací u rostlin).

2. Cílené změny jsou záležitostí celé baterie proteinů – enzymů, které je způsobují. Například k fosforylaci serinu potřebujeme enzym protein kinázu. A protože jde o enzymy, ty rozpoznávají nejen cílovou aminokyselinu, tj. serin S, ale také tvar jejího okolí: může se stát, že bude třeba jedna kináza k fosforylaci serinu v poloze 10 (okolí – epitop RKST), jiná pro polohu 28 (RKSF) a ještě jiná pro epitop RµSF; podobně pro enzymy – metylázy lysinů 4, 9 a 23, s tím, že pro produkci trimetyllysinu jsou třeba enzymy tři – pro každý stupeň metylace jiný. Z celé baterie modifikujících enzymů může mít buňka v daném místě a čase k dispozici jen některé, ostatní sice jsou v pohotovosti, ale v dané chvíli neúčinné, a další se budou syntetizovat dle potřeby. Také nutno uvést, že příslušný enzym „neútočí“ na všechny proteiny vlastnící daný epitop, ale jen na vybranou množinu, ke které má přístup.
3. Obrovskou výhodou epimutací je také to, že jsou vratné: například účinky kinázy na serin lze zrušit fosfatázou fosfoserinu (opět více nebo méně citlivou pro jistý epitop), metylaci lysinu ruší demetyláza apod.
Modelový příklad: Enzym E1 katalyzuje látku L, enzym E2 ji rozkládá. E1 je aktivní, E2neaktivní, L se hromadí. Potřebuje-li buňka L naopak rozkládat, „pošle“ kinázu, která nafosforyluje oba enzymy (znázorněno jako P). Tím změní jejich konformaci, a to tak, že E1Pbude vypnut, E2P se aktivuje. Tento režim bude trvat do té doby, než příslušná fosfatáza opět vrátí oba enzymy do předchozího stavu. Protože na jeden protein lze „naložit“ celou řadu epimutací různého druhu, lze jeho funkci nastavovat nejen v předvedeném režimu „vypni / zapni“, ale také tuto funkci dle potřeby ladit (viz níže histony). Většina, snad 90 % druhů proteinů přítomných v buňce jsou proteiny sítě regulačních drah, a ty různou derivatizací umožňují přenos nebo jemné ladění signálů – často tak, že po příslušné úpravě se celá řada (často desítky) různých proteinů shluknou a tato částice pak přenáší potřebný signál. Životnost částice bývá krátká, odstraněním některé klíčové epimutace se komplex opět rozpadá. Níže si předvedeme jiný druh shlukování proteinů, které v jaderném chromatinu regulují expresi genů.
4. Srovnejme derivatizaci proteinů s epigenetickými mutacemi DNA (viz Aleš Kovařík: Metylace cytosinu jako přidaná hodnota DNA, Vesmír 2/2015). Tam je omezení víc. V první řadě úpravy DNA nesmí překážet její replikaci ani opravám: komplex DNA polymerázy i opravných enzymů musí mít přístup k dvoušroubovnici a chemické modifikace bází nerozpoznává (jako kdyz jsme jeste nedavno mailovali bez hacku a carek). Výhodou však je jednoduchý způsob, jak se na dceřiné vlákno – podle mateřského – s pomocí příslušných enzymů dle potřeby kopírují nebo přestanou kopírovat některé epimutace. Výhodou je také přenesení úprav do další generace buněk či někdy dokonce organismů. (Viz Vít Latzel: Epigenetická dědičnost v ekologii a evoluci rostlin, Vesmír 2/2015; Vojtěch Čermák, Šárka Motylová: Džungle epigenetických modifikací u rostlin.)
**
Chromatin
Pokud jde o epigenetické procesy u eukaryotních buněk, nemůžeme opomenout histony a jejich roli při manipulaci buňky s vlastní DNA. Histony jsou evolučně velmi konzervativní proteiny, na kterých je v jádře namotána nositelka dědičnosti, deoxyribonukleová kyselina DNA – představme si to zhruba jako nit namotanou na „špulce“, kdy je každá jedna špulka (říká se jí nukleozom) tvořena osmi histonovými proteiny (čtyři dvojice) a devátý histon pak ještě slouží jako speciální spojka mezi špulkou a DNA řetězci. Nukleozomy nejenže DNA pomáhají kondenzovat v jádře, když se mnohonásobně delší DNA molekula musí vměstnat do jádra o velikosti kolem 10 mikrometrů, ale pomáhají i při regulaci přepisu genů, opravě DNA či rekombinaci (obr. 2).
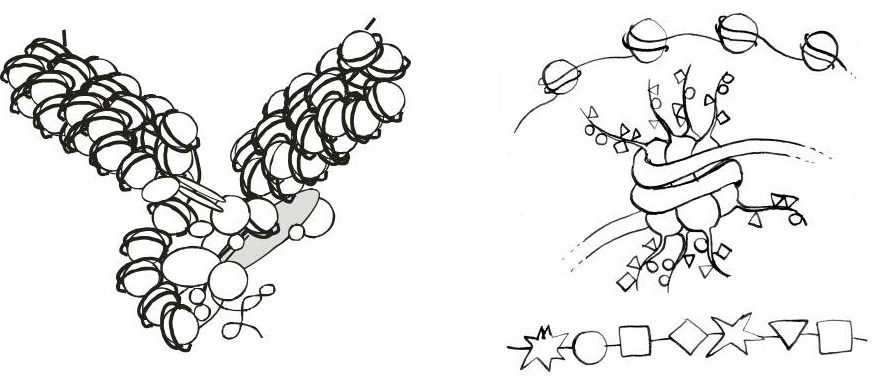
Histony jsou na N-koncích svých řetězců za pomoci velmi specifických proteinů chemicky modifikovány, což znamená, že jejich aminokyseliny (většinou jde o lysiny, argininy či seriny) mohou být acetylovány, metylovány (jedenkrát až třikrát), fosforylovány či označeny jinými „značkami“, jako je protein ubiquitin nebo cukr ribóza. To, že jsou histony takto modifikovány, je známo již od roku 1964 především díky výzkumu Vincenta Allfreyho, který je považován za otce epigenetiky (samotné objevení histonů Albrechtem Kosselem se datuje již o století dříve); na to, abychom mohli zjistit, jak přesně tyto modifikace ovlivňují chromatin (to znamená nejen DNA, ale i všechny proteiny a RNA molekuly, které jsou na DNA v jádře navázány), jsme museli čekat skoro až do konce minulého století, kdy se zdokonalily příslušné metody zkoumání.
Zmiňované modifikace histonových aminokyselin mohou ovlivňovat přímo strukturu chromatinu tak, že mění jeho náboj a ruší elektrostatické interakce mezi DNA a histonovými proteiny, což vede k rozvolnění DNA z histonů a zpřístupnění genetického zápisu. V těchto případech jde většinou o acetylaci a fosforylaci, ale někdy má přítomnost těchto značek přesně opačnou funkci, tj. kondenzaci chromatinu a znemožnění přepisu genů. Velmi často totiž záleží na kombinatorice modifikací, na tom, na jakém místě se přesně jaká značka v aminokyselinovém řetězci nachází a zda jde o jednu, dvě či trojitou metylaci (nebo o jinou značku) či kombinaci značek. Histonové modifikace totiž slouží především jako vazebný substrát pro další proteiny, které je dokážou „přečíst“ a navázat. Tyto vazebné proteiny pak za spotřeby energie rozvolňují DNA namotanou na histonech a zpřístupňují genetický zápis přepisu či opravě, nebo naopak semknou DNA řetězce a histonové proteiny tak, že se DNA stane zcela nepřístupnou (a umlčenou), a to někdy i po celý život svého nositele.
Modifikace nejsou trvalé a transkripce genů může být dynamicky kontrolována neustálými změnami „diakritiky“ histonových aminokyselin – v odpovědi na signály z prostředí i vnitřní stav buňky. Pochopitelně se vše odehrává v jednotě s modifikacemi DNA a množinou přítomných typů RNA. Modifikace na histonech tvoří jakýsi čárový kód vyšší úrovně: například u člověka teoreticky až 3 miliony různých nukleozomových variant, kdy každá nese jinak uspořádané modifikace. Díky této obrovské varietě pak mohou vznikat jednotlivé buněčné typy, kdy specializované buňky srdce, jater, mozku a dalších orgánů nesou (více či méně) shodný genetický zápis, ale teprve rozdíly v tom, jaké geny jsou a nejsou v daném buněčném jádře aktivní, stojí za konkrétní podobou buněk.
Celý tento histonový „čárový kód“ je určen ekosystému stovek proteinů, které rozpoznávají příslušné histony a rozhodují o tom, které části genetické informace budou v daném kontextu rozpoznávány a exprimovány.
O autorech
Jana Švorcová
Anton Markoš
Další články k tématu
Džungle epigenetických modifikací u rostlin
Čeká nás století kmenových buněk?
Střevní bakterie řídí aktivitu lidských genů
Jak se dělá potomek: Aristotelés o rozmnožování živočichů
Jak potrava ovlivňuje vzhled hmyzu
Malé RNA: Mocní trpaslíci
Malá historie epigenetiky
Úvodní slovo - všechno je epigenetika?
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
